4偮偺ZIP僞儞僷僋幙偑丄崻偺奜懁椞堟偵偍偗傞Zn丄Fe丄Cu丄偍傛傃Mn偺媧廂偵婑梌偡傞
傾儔價僪僾僔僗偺Zip family偺堚揱巕偺偆偪丄Zip2,Zip3,Zip5,Zip8偺徻嵶側夝愅傪峴偭偰丄垷墧傗摵埲奜偵揝媧廂偵娭楢偡傞堚揱巕Zip2,Zip8傪尒弌偟偨曬崘偱偁傞丅巻柺偺娭學忋夋憸偱偼丄恾2丄恾3丄恾5偺傒傪徯夘偟偨丅
4偮偺ZIP僞儞僷僋幙偑丄崻偺奜懁椞堟偵偍偗傞Zn丄Fe丄Cu丄偍傛傃Mn偺媧廂偵婑梌偡傞
Four ZIPs contribute to Zn, Fe, Cu and Mn acquisition at the outer root domain
働價儞丒儘乕僽1丄儕儞僇丒儖僼僃僽儖亖儗僕僃儞僪儖1丄僼傽價僄儞僰丒僋儗乕儖1丄儅儕乕丒僶儖儀儘儞1
僕儏僱乕僽戝妛 怉暔壢妛晹栧丄30 僋傾僀丒僄儖僱僗僩丒傾儞僙儖儊丄1211 僕儏僱乕僽丄僗僀僗
bioRxiv preprint doi: https://doi.org/10.1101/2024.10.31.621270乮枹庴棟榑暥乯
梫栺
垷墧乮Zn乯偼怉暔偺惉挿偵晄壜寚側旝検尦慺偱偁傝丄怉暔偺敪払偵廳梫側栶妱傪壥偨偟傑偡丅偟偐偟丄憃巕梩怉暔偵偍偗傞搚忞偐傜偺垷墧媧廂偵娭梌偡傞嬶懱揑側桝憲懱偼傑偩柧妋偵側偭偰偄傑偣傫丅Arabidopsis thaliana偺promoter-reporter儔僀儞傪梡偄偰丄昞旂偱敪尰偟丄奜崻椞堟偐傜偺Zn媧廂偵娭梌偡傞壜擻惈偺偁傞ZIP乮Zn挷愡桝憲懱丄揝挷愡桝憲懱乮IRT乯條僞儞僷僋幙乯僼傽儈儕乕儊儞僶乕傪暋悢摨掕偟傑偟偨丅ZIP2丄ZIP3丄ZIP5丄偍傛傃ZIP8偼丄昞旂嵶朎偲旂憌嵶朎偺嵶朎枌偵庡偵嬊嵼偟丄搚忞偐傜偺嬥懏媧廂偵偍偗傞愽嵼揑側栶妱傪巟帩偟偰偄傑偡丅惗棟妛揑尋媶丄僀僆儞僾儘僼傽僀儕儞僌丄偍傛傃堚揱妛揑夝愅傪捠偠偰丄ZIP3偲ZIP5偑Zn媧廂偺庡梫側峷專幰偱偁傞偙偲偑敾柧偟丄堦曽丄ZIP2偲ZIP8偼偦傟偧傟摵乮Cu乯偲揝乮Fe乯偺媧廂偵庡偵娭梌偟偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丅摿偵丄ZIP3偲ZIP8偼崻偺昞旂嵶朎偵偍偄偰IRT1偲摨條偺奜懁嬌惈傪帵偟丄桝憲懱偺嬌惈偑峼暔媧廂偵廳梫側栶妱傪壥偨偡偙偲傪嫮挷偟偰偄傑偡丅偙傟傜偺敪尒偼丄怉暔崻偵偍偗傞嬥懏媧廂偺儊僇僯僘儉偵娭偡傞怴偨側摯嶡傪採嫙偟丄怉暔偺嬥懏娷桳検傪岦忋偝偣傞偨傔偺僶僀僆僼僅乕僥傿僼傿働乕僔儑儞愴棯偺壜擻惈傪帵嵈偟偰偄傑偡丅
恾偺杴椺
恾1. 垷墧寚朢偑怉暔偺惉挿偲僀僆儞慻惉偵梌偊傞塭嬁
乮A乯 垷墧廩懌忦審乮+Zn乯偲垷墧寚朢忦審乮-Zn乯偱3廡娫攟梴偟偨栰惗宆乮WT乯怉暔偺戙昞揑側夋憸丅悈暯僾儗乕僩乮忋乯偲悅捈僾儗乕僩乮壓乯偵昞帵丅僗働乕儖僶乕 : 1 cm丅僶僀僆儕儞僾儘僢僩偼丄乮B乯慛廳乮FW乯丄乮C乯僋儘儘僼傿儖娷検丄偍傛傃乮D乯Fv/Fm傪帵偡丅埫弴墳偝偣偨WT怉暔傪垷墧廩懌忦審乮+Zn丄惵乯傑偨偼垷墧寚朢忦審乮-Zn丄僆儗儞僕乯偱3廡娫攟梴偟偨寢壥丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪昞偡乮n≥10乯丅乮E乯宻偲乮F乯崻偺僀僆僲乕儉僾儘僼傽僀儖丅栰惗宆怉暔傪垷墧廩懌忦審乮+Zn乯傑偨偼垷墧寚朢忦審乮-Zn乯壓偱3廡娫攟梴偟偨寢壥丅僨乕僞偼+Zn忦審傪100偲偟偰憡懳抣偱帵偝傟丄3 撈棫偟偨僾乕儖偐傜摼傜傟偨暯嬒抣偵懳墳偡傞丅
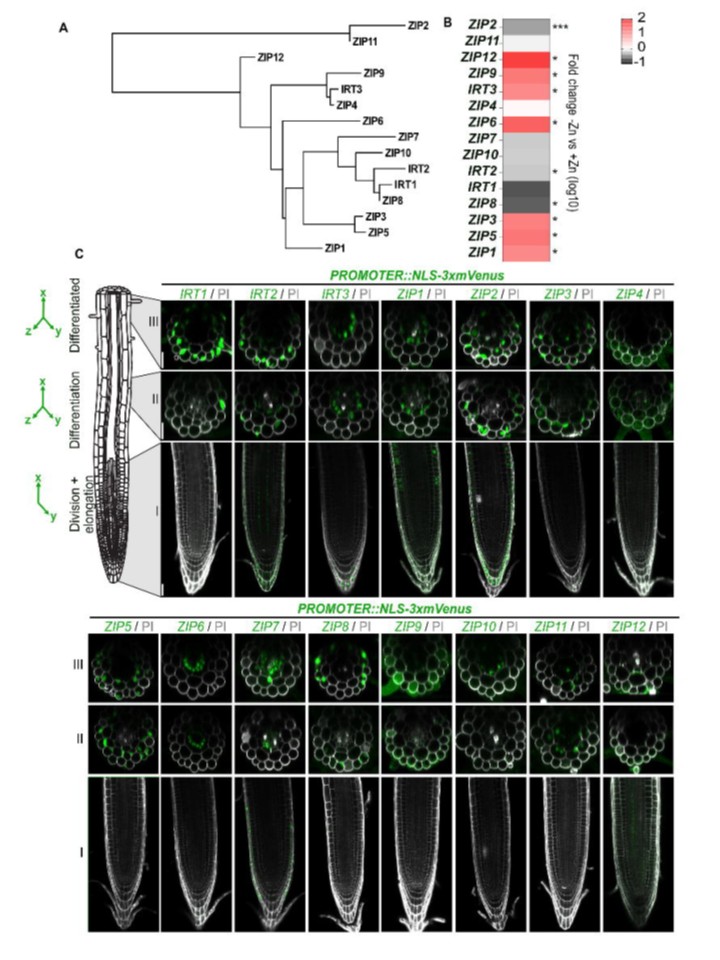
恾2. Arabidopsis thaliana偺崻偵偍偗傞AtZIP僼傽儈儕乕偺敪尰偺嬻娫揑丒帪娫揑僷僞乕儞 塱懕惈丅
(A) 傾儔價僪僾僔僗丒僞儕傾僫乮Arabidopsis thaliana乯偺AtZIP僼傽儈儕乕15儊儞僶乕偺宯摑庽丅偙偺宯摑庽偼丄Phylogeny.fr傪巊梡偟偰僞儞僷僋幙攝楍偐傜惗惉偝傟偨丅
(B) 惂屼忦審壓乮+Zn乯傑偨偼Zn寚朢忦審壓乮-Zn乯偱1廡娫攟梴偝傟偨栰惗宆怉暔偺崻偵偍偗傞15偺ZIP儊儞僶乕偺憡懳揑揮幨検丅寢壥偼僸乕僩儅僢僾偱帵偝傟丄惂屼忦審乮+Zn乯偲偺斾妑偱偺攞憹曄壔偑嫮挷偝傟偰偄傞丅僨乕僞偼4偮偺惗暔妛揑暋惢偐傜摼傜傟丄奺暋惢偼彮側偔偲傕40杮偺崻偺僾乕儖偐傜峔惉偝傟偰偄傞丅
(C) 堎側傞崻偺椞堟偵偍偗傞ZIP::NLS-3xmVenus偺敪尰僷僞乕儞傪帵偡戙昞揑側夋憸丗椞堟I乮崻偺愭抂丄壓抜偺夋憸乯丄椞堟II乮憗婜暘壔椞堟丄拞抜偺夋憸乯丄椞堟III乮惉弉椞堟丄忋抜偺夋憸乯丅
妀嬊嵼惈偺mVenus僔僌僫儖偼椢怓偱帵偝傟丄嵶朎暻傪愼怓偡傞偨傔偺PI偼奃怓偱帵偝傟偰偄傞丅夋憸偼崻偵増偭偰奺儔僀儞偱摨堦偺愝掕偱嶣塭偝傟偨偑丄ZIP娫偺僾儘儌乕僞乕妶惈嵎偺偨傔丄儔僀儞娫偱愝掕偑堎側偭偨丅僝乕儞I偱偼丄mVenus偲PI偺扨堦徟揰夋憸乮XY乯偑昞帵偝傟丄僝乕儞II偲III偱偼丄Z-僗僞僢僋偺嵟戝搳塭乮XYZ乯偲偟偰mVenus僔僌僫儖偑昞帵偝傟丄Z-僗僞僢僋偐傜拪弌偝傟偨PI偺扨堦捈岎價儏乕偑廳偹傜傟偰偄傞丅夋憸偼T1怉暔偵懳墳偡傞丅彮側偔偲傕5偮偺撈棫偟偨T1怉暔偑夋憸壔偝傟偨丅僔僌僫儖偼T2怉暔偱傕妋擣偝傟偨丅僗働乕儖僶乕丗偡傋偰偺夋憸偱25 兪m丅
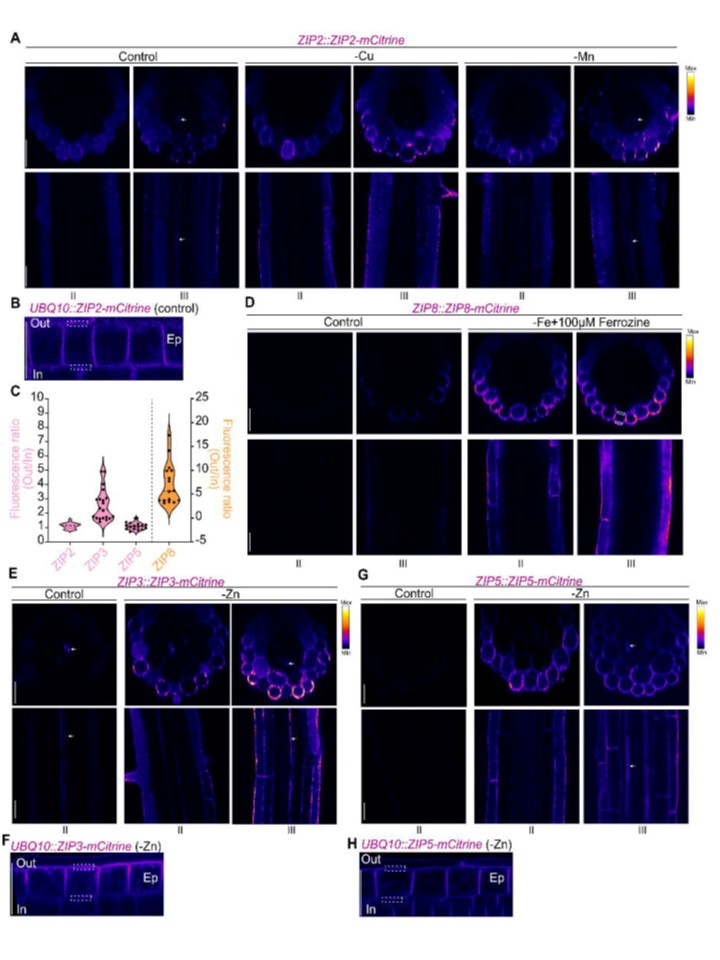
恾3. ZIP2丄ZIP8丄ZIP3偍傛傃ZIP5偺奜崻椞堟偵偍偗傞嬊嵼壔
(A丄D丄E偍傛傃G) ZIP2丄ZIP8丄ZIP3傑偨偼ZIP5傪撪嵼惈僾儘儌乕僞乕偺惂屼壓偱mCitrine偲梈崌偝偣偨5擔楊偺昪偵偍偗傞mCitrine寀岝傪帵偡戙昞揑側嫟徟揰夋憸丅昪偼丄僐儞僩儘乕儖忦審傑偨偼嬥懏寚朢忦審乮-Cu丄-Mn丄-Fe + 100 兪M Ferrozine丄-Zn乯偱攟梴偝傟偨丅忋抜偺僷僱儖偼崻偺墶抐柺夋憸乮Z僗僞僢僋偐傜拪弌乯傪帵偟丄壓抜偺僷僱儖偼崻偺拞墰徟揰暯柺偱嶣塭偝傟偨廲抐柺夋憸傪帵偡丅懳墳偡傞崻偺椞堟乮恾2C偱掕媊偝傟偨II傑偨偼III乯偼夋憸偺壓晹偵帵偝傟偰偄傞丅偡傋偰偺夋憸偼T2怉暔偐傜庢摼偝傟偨丅(A, B, D-H) mCitrine偺寀岝偼Fire儖僢僋傾僢僾僥乕僽儖乮LUT乯傪巊梡偟偰昞帵偝傟偰偄傞丅(B, F, H) 5擔楊偺昪偱UBQ10::ZIP2/3/5-mCitrine傪敪尰偟丄僐儞僩儘乕儖乮ZIP2乯傑偨偼-Zn忦審乮ZIP3偲ZIP5乯壓偱攟梴偝傟偨怉暔偺mCitrine寀岝僔僌僫儖偺嫟徟揰夋憸丅崻偼怢挿椞堟偺昞旂乮Ep乯嵶朎偺拞墰徟揰暯柺偱夋憸壔偝傟丄嬌惈嬊嵼傪昡壙偟偨丅(C) 僷僱儖B丄F丄H乮僺儞僋乯偵帵偝傟偨ZIP2丄ZIP3丄偍傛傃ZIP5-mCitrine偺寀岝斾暘愅丅ZIP8-mCitrine斾乮僆儗儞僕乯偼丄崻偺僝乕儞III偺昞旂偱寁嶼偝傟偨丅寀岝斾偼丄奺昞旂嵶朎偺奜懁乮Out乯PM偺暯嬒寀岝嫮搙傪撪懁乮In乯PM偺暯嬒寀岝嫮搙偱妱傞偙偲偱寛掕偝傟偨丅偙偺嫮搙斾偼丄奺昞旂嵶朎偺奜懁乮Out乯PM偺嫮搙偲撪懁乮In乯PM偺嫮搙偵婎偯偄偰嶼弌偝傟偨丅奜懁偲撪懁偺寀岝掕検偵偼摨偠椞堟乮ROI乯偑巊梡偝傟偨丅Out/In斾傪掕検偡傞偨傔偵巊梡偝傟偨ROI偺椺偼丄奺堚揱巕宆乮恾B丄D丄F丄H乯偛偲偵攋慄巐妏宍偱帵偝傟偰偄傞丅斾棪偼丄奺崻偵偮偒彮側偔偲傕5偮偺嵶朎偲丄彮側偔偲傕3偮偺撈棫偟偨崻偐傜寁嶼偝傟偨丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣傪昞偟丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪昞偟傑偡乮n≥15乯丅僗働乕儖僶乕丗偡傋偰偺夋憸偱25 兪m丅
恾4. zip2曄堎懱偵偍偗傞摵偺拁愊忈奞
(A) 惂屼忦審壓乮+Cu乯偲摵寚朢忦審壓乮-Cu乯偱10擔娫悅捈攟梴偟偨栰惗宆乮WT乯偲2偮偺zip2曄堎懱傾儗儖偺戙昞揑側夋憸丅奺堚揱巕宆偐傜3偮偺戙昞揑側怉暔傪傾僈乕僾儗乕僩偵堏偟丄夋憸壔傪峴偭偨丅僗働乕儖僶乕丗1 cm丅 (B-D) +Cu傑偨偼-Cu忦審壓偱10擔娫攟梴偟偨怉暔偺(B)慛廳(FW)丄(C)崻挿丄偍傛傃(D)僋儘儘僼傿儖娷桳検偺暘晍傪帵偡僶僀僆儕儞僾儘僢僩乮B偲C偼n亜15丄D偼n亜5乯丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣傪昞偟丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪帵偡丅乮E乯僐儞僩儘乕儖傾僈乕僾儗乕僩忋偱3廡娫悅捈偵攟梴偝傟偨WT偲zip2-1曄堎懱偺宻乮嵍乯偲崻乮塃乯偵偍偗傞嬥懏娷検乮Fe丄Cu丄Mn丄Zn乯丅奺惗暔妛揑暋惢偛偲偵3乣4姅傪僾乕儖偟偨乮n=4乯丅
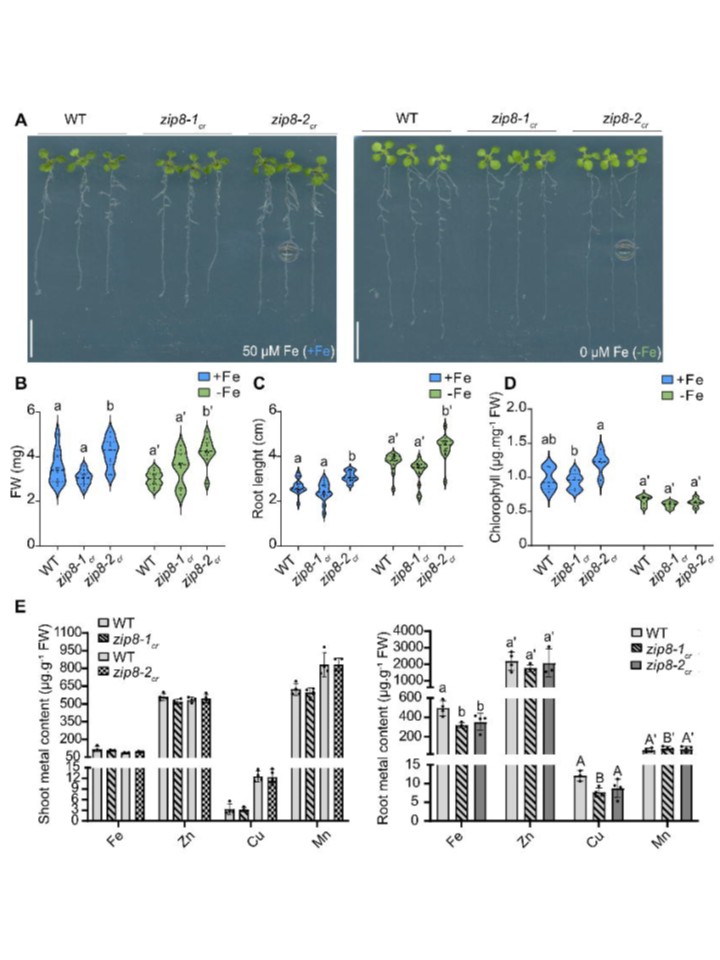
恾5. zip8曄堎懱偵偍偗傞揝偺拁愊忈奞
(A) 惂屼忦審壓乮+Fe乯偲揝寚朢忦審壓乮-Fe乯偱10擔娫悅捈攟梴偟偨栰惗宆乮WT乯偲2偮偺zip8曄堎懱傾儗儖偺戙昞揑側夋憸丅奺堚揱巕宆偵偮偒3姅傪傾僈乕僾儗乕僩偵堏偟丄夋憸壔傪峴偭偨丅僗働乕儖僶乕丗1cm丅 (B-D) 恾A偱愢柧偝傟偨忦審壓偱攟梴偝傟偨怉暔偺(B) 慛廳丄(C) 崻挿丄偍傛傃(D) 僋儘儘僼傿儖娷桳検偺暘晍傪帵偡償傽僀僆儕儞僾儘僢僩乮B偲C偼n亜15丄D偼n亜5乯丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣傪昞偟丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪帵偟傑偡丅堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅乮ANOVA乯偵懕偄偰Tukey's偺帠屻専掕傑偨偼僋儖僗僇乕儖丒僂僅儕僗専掕偵懕偄偰僟儞儞偺専掕乮P < 0.05乯偵傛傝摑寁揑偵桳堄側嵎傪帵偡偙偲傪帵偡丅 (E) 惂屼忦審壓偱3廡娫悅捈偵攟梴偝傟偨WT偲zip8曄堎懱偺宻乮嵍乯偲崻乮塃乯偵偍偗傞嬥懏娷桳検乮Fe丄Cu丄Mn丄Zn乯丅奺惗暔妛揑暋惢偱偼丄3乣4姅傪僾乕儖偟偨乮n=4乯丅僨乕僞偼暯嬒 亇 昗弨岆嵎偱帵偝傟丄宻偺僨乕僞偱偼桳堄嵎偑擣傔傜傟側偐偭偨偨傔丄Student偺t専掕傑偨偼Mann-Whitney専掕偱堚揱巕宆娫偺桳堄嵎傪敾掕偟偨丅堦曽丄崻偺僨乕僞偼堦尦攝抲暘嶶暘愅乮ANOVA乯偵懕偄偰Tukey偺帠屻専掕傪幚巤偟偨乮P < 0.05乯丅
恾6. ZIP3偍傛傃ZIP5偵傛傞垷墧偺媧廂
(A) 懳徠忦審乮+Zn乯偍傛傃垷墧寚朢忦審乮-Zn乯壓偱10擔娫悅捈偵攟梴偝傟偨栰惗宆怉暔丄zip3-1丄zip5-1丄偍傛傃擇廳曄堎懱zip3-1zip5-1偺戙昞揑側夋憸丅奺堚揱巕宆偺戙昞揑側3偮偺怉暔傪傾僈乕僾儗乕僩偵堏偟丄夋憸壔傪峴偭偨丅僗働乕儖僶乕丗1cm丅 (B-D) 恾A偱愢柧偝傟偨忦審壓偱攟梴偝傟偨怉暔偺(B) 慛廳丄(C) 崻挿丄偍傛傃(D) 僋儘儘僼傿儖娷桳検偺暘晍傪帵偡償傽僀僆儕儞僾儘僢僩乮B偲C偼亜15丄D偼n亜5乯丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣傪昞偟丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪帵偡丅
堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅乮ANOVA乯偵懕偄偰Tukey偺帠屻専掕傑偨偼Kruskal乕Wallis専掕偵懕偄偰Dunn偺専掕偵傛傝摑寁揑偵桳堄側嵎傪帵偡偙偲傪帵偡乮P < 0.05乯丅(E) WT偲zip3-1zip5-1曄堎懱偺3廡娫攟梴偟偨宻乮嵍乯偲崻乮塃乯偺嬥懏娷検乮Fe丄Cu丄Mn丄Zn乯丅奺惗暔妛揑暋惢偛偲偵丄3乣4姅傪僾乕儖偟偨乮n=4乯丅(F) 5擔楊偺WT偲zip3-1zip5-1崻偺戙昞揑側嫟徟揰夋憸丅20 兪M Zinpyr-1偱3帪娫愼怓偟偨丅愼怓慜偵丄怉暔偼5擔娫Zn寚朢忦審壓偱攟梴偝傟偨丅忋抜偼抐柺恾丄壓抜偼暘壔偟偨崻偺嵟戝搳塭恾傪帵偡丅僗働乕儖僶乕丗25兪m丅忋抜偺僷僱儖偼抐柺恾傪帵偟丄壓抜偺僷僱儖偼暘壔偟偨崻偺嵟戝搳塭恾傪帵偟偰偄傞丅僗働乕儖僶乕丗25兪m丅 (G) WT偲zip3-1zip5-1崻偺娭怱椞堟乮ROI乯偵偍偗傞撪泱梩偺Zinpyr-1僔僌僫儖偺暘晍傪帵偡僶僀僆儕儞僾儘僢僩丅彮側偔偲傕4偮偺撈棫偟偨崻偑暘愅偝傟偨乮n亜15乯丅償傽僀僆儕儞僾儘僢僩偺攋慄偼拞墰抣丄揰慄偼戞1巐暘埵悢偲戞3巐暘埵悢傪昞偟傑偡丅乮H乯僐儞僩儘乕儖傾僈乕僾儗乕僩忋偱3廡娫攟梴偝傟偨WT丄ZIP3丄ZIP5夁忚敪尰宯摑偺崻偵偍偗傞嬥懏娷桳検乮Fe丄Cu丄Mn丄Zn乯丅奺惗暔妛揑暋惢偵偍偄偰丄3乣4偮偺T1怉暔傪僾乕儖偟偨乮n=4乯丅僨乕僞偼暯嬒 亇 SE 偱帵偝傟丄堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅乮ANOVA乯 followed by Tukey偺帠屻専掕乮P < 0.05乯偵傛傝摑寁揑偵桳堄側嵎傪帵偡丅(I) 5擔楊偺WT丄35S::ZIP3乮T1乯偍傛傃35S::ZIP5乮T1乯怉暔偺戙昞揑側嫟徟揰夋憸丅20 兪M Zinpyr-1偱3帪娫愼怓偟偨丅愼怓慜偵1 兪M Zn偱5擔娫攟梴偟偨丅忋抜偺僷僱儖偼暘壔偟偨崻偺抐柺傪帵偟乮僗働乕儖僶乕丗25 兪m乯丄壓抜偺僷僱儖偼崻偺愭抂偐傜偺嵟戝搳塭傪帵偟傑偡乮僗働乕儖僶乕丗200 兪m乯丅
恾7. ZIP3偺嬊嵼壔偼垷墧偺壜梡惈偵埶懚偡傞
(A-G) UBQ10::ZIP3-mCitrine 庬巕昪傪Zn乮15 兪M乯傪娷傓傾僈乕僾儗乕僩忋偱5擔娫攟梴偟丄偦偺屻丄Zn寚朢忦審乮-Zn乯傑偨偼Zn偺10攞夁忚検乮10x Zn丟150 兪M乯壓偱16帪娫塼懱攟抧偵堏偟丄夋憸壔慜偵攟梴偟偨丅(A) UBQ10::ZIP3-mCitrine傪敪尰偡傞5擔楊偺崻偺戙昞揑側嵟戝搳塭夋憸丅mCitrine僔僌僫儖偼墿怓偱帵偝傟偰偄傞丅僗働乕儖僶乕丗125 兪m丅 (B) 恾A偵帵偡暋悢偺撈棫偟偨崻偺嵟戝搳塭夋憸偐傜丄ZIP3-mCitrine僔僌僫儖嫮搙傪掕検壔偟偨寢壥乮n亜5乯丅僨乕僞偼暯嬒亇昗弨曃嵎偱帵偝傟偰偄傞丅儅儞丒儂僀僢僩僯乕専掕乮P < 0.05乯偵傛傝丄忦審娫偺桳堄側嵎偼娤嶡偝傟側偐偭偨丅
(C, D) 僷僱儖A偵帵偝傟偨崻偺怢挿椞堟偺奼戝恾丅夋憸偼昞旂偺恀傫拞偺徟揰暯柺乮C乯偲昞旂偺昞柺乮D乯偱嶣塭偝傟偨丅mCitrine僔僌僫儖偼Fire乮LUT乯僇儔乕僗働乕儖偱昞帵偝傟偰偄傞丅僗働乕儖僶乕丗12.5 兪m丅 (E) ZIP3-mCitrine僔僌僫儖偺嵶朎枌乮PM乯偲嵶朎撪乮intra乯偺斾棪偺掕検壔乮n亜20乯丅僨乕僞偼暯嬒 亇 昗弨曃嵎偱帵偝傟丄忦審娫偺摑寁揑嵎堎偼儅儞丒儂僀僢僩僯乕専掕偱寛掕偝傟偨乮*P < 0.05丟**P < 0.01丟 ***P < 0.0001乯丅(F) 忋婰偺曽朄偱攟梴偝傟偨怉暔偺昞旂嵶朎偺昞柺憸丅夋憸壔慜偵100 兪M CHX偲50 兪M BFA偵敇業偝傟偨傕偺丅mCitrine僔僌僫儖偼Fire (LUT) 僇儔乕僗働乕儖偱昞帵偝傟偰偄傞丅僗働乕儖僶乕丗25 兪m丅(G) 僷僱儖F偵婰嵹偺曽朄偱張棟偝傟偨怉暔偵偍偗傞ZIP3-mCitrine僔僌僫儖偺PM懳嵶朎撪乮intra乯斾偺掕検壔乮n亜20乯丅僨乕僞偼暯嬒亇昗弨曃嵎偱帵偝傟丄忦審娫偺摑寁揑嵎堎偼Mann-Whitney専掕偱寛掕偝傟偨乮*P < 0.05丟**P < 0.01丟***P < 0.0001乯丅
恾2
恾俁
恾俆