(重要論文)キレート化に基づく鉄吸収は、冷地型牧草における長期高温ストレスの影響を緩和する
この論文は温暖化のせいで通常の気候よりも高温が長く続く場合に、小麦が鉄欠乏クロロシスを起こす。その結果収量減となる。その際に野副朋子さん(現・乙津朋子)が発見したムギネ酸排出トランスポーターである「TOM1系遺伝子」が、高温耐性品種で、根や地上部で強く発現していることを見出したものである。
また、愛知製鋼の鈴木基史君が提供した人工合成ムギネ酸誘導体である「PDMA」を根圏に投与すると、この高温クロロシスが顕著に回復するので、生体内でのムギネ酸合成能と排出能が、高温耐性に必須であることを明らかにしたものである。
小生(森敏)は10年以上前にベゴニアの激しい高温誘導クロロシスが鉄欠乏によるものであると予想し、そのことを winep blog
2010/10/03 : 気候が涼しくなって、回復しはじめたベゴニアのクロロシス症状
http://moribin.blog114.fc2.com/blog-entry-915.html
2010/09/03 : ベゴニアのクロロシス
http://moribin.blog114.fc2.com/blog-entry-900.html
で紹介しておいた。この nature communications の論文で、小生の予想「高温誘導性鉄欠乏が存在する」という仮説が証明されたのである。
また、この論文は、将来の温暖化気候下での作物栽培に対して、国産のPDMAが葉面散布などに有効利用されうる道を開いたものである。
キレート化に基づく鉄吸収は、冷地型牧草における長期高温ストレスの影響を緩和する。
Chelation-based iron uptake mitigates the effects of prolonged high-temperature stress in cool-season grasses
Anzu Minami, Yoshihiko Onda, Minami Shimizu, Yukiko Uehara-Yamaguchi, Asaka Kanatani, Risa Nakayama, Kyoko Toyama, Kotaro Takahagi, Komaki Inoue, Tomoko Nozoye, Motofumi Suzuki, Yusuke Kouzai, Toshihisa Nomura, Keitaro Tanoi, Masaki Endo, Ryuji Miki, Masakazu Kashihara, Naoaki Taoka & Keiichi Mochida
Nature Communications volume 16, Article number: 7709 (2025)
要旨
高温ストレスは、農業生産性と生態系多様性に重大な脅威をもたらします。本研究では、長期にわたる高温ストレスがコムギ(Triticum aestivum)とモデルイネ科植物Brachypodium distachyonに及ぼす影響を示し、熱ストレスが新葉の鉄欠乏を引き起こすことを実証します。B. distachyonの量的形質遺伝子座位解析により、ムギネ酸ファミリーフィトシデロフォア輸送体1遺伝子(BdTOM1)を含む、耐熱性と関連するゲノム領域が明らかになりました。BdTOM1を含む鉄欠乏関連遺伝子は、高温耐性B. distachyon系統において、感受性系統よりも高温時に高発現し、その結果、デオキシムギネ酸の分泌量が増加します。プロリン-2'-デオキシムギネ酸の処理は熱誘導性生育阻害を軽減しますが、過剰な鉄処理は両種において毒性を引き起こします。本研究の結果は、長期にわたる高温ストレスにおける熱誘導性栄養ストレスの役割を浮き彫りにし、鉄の恒常性が極端な気候に対する作物の耐性を向上させるための有望なターゲットとなり得ることを示唆しています。
はじめに
気候変動は植物の生産性と作物の収量を脅かしており、特に小麦(Triticum aestivum)のような冷涼期穀物において顕著である。これらの作物は、生育期および生殖期における熱ストレスへの曝露増加により、大幅な収量損失が生じると予測されている。熱ストレスは光合成や呼吸といった重要な生理反応を阻害し、収量低下を招く。植物細胞内では、熱ストレスが原形質膜の流動性を高め、葉緑体やミトコンドリアの電子伝達系を損なうことで、活性酸素種(ROS)の過剰産生、タンパク質の誤折り畳み、脂質過酸化、DNA損傷、細胞内恒常性の破壊を引き起こす。これらの悪影響を軽減するため、植物は熱ショック応答を通じて熱ストレスを感知し対応する。これにはカルシウムシグナル経路の活性化、膜完全性の維持、細胞死の防止、熱ショック因子(HSF)遺伝子の転写が含まれる。HSF遺伝子は、ROS除去に関与し分子シャペロン機能を有する熱ショックタンパク質(HSP)遺伝子の発現を調節するタンパク質をコードする。植物の熱ストレス応答を探る実験設計は、主に最適生育温度より15℃以上高い温度への曝露を用いた短期的な熱ショック処理に基づいている。しかし気候変動は、中程度の高温(約35℃)での長期的な熱ストレスをもたらし、生育からストレス順化への持続的な移行を引き起こす。これにより植物の発育、成長、生殖が遅延する。長期にわたる中程度の熱ストレスに対する植物の反応を理解することは、気候変動に対する作物の回復力を向上させる上で極めて重要である。
鉄は光合成と呼吸において重要な役割を担う必須微量栄養素である。植物細胞内の鉄濃度は、鉄過剰と鉄欠乏の両方を回避するために厳密に調節されなければならない。土壌中の鉄は豊富に存在するが、主に不溶性の三価鉄(Fe3+)として存在するため、その生物学的利用能は限られている。したがって、イネ(Oryza sativa)、小麦、ブラキポディウム・ディスタキオンなどのイネ科植物は、主にキレート化に基づく戦略IIメカニズムに依存して土壌からFe3+を吸収する。これらの植物はムギネ酸などの植物性鉄輸送体を生成し、ムギネ酸ファミリー植物性鉄輸送体(TOM)様排出輸送体を介して根圏に放出され、Fe3+をキレート化する。
Fe3+-フィトシデロフォア複合体は、YELLOW STRIPE 1およびYELLOW STRIPE1-LIKE(YS1、YSL)ファミリー成員などの鉄輸送体を介して細胞内に取り込まれる。しかし、中程度の熱ストレスがイネ科植物の鉄恒常性を損なうかどうかは不明である。
本研究では、パン小麦とB. distachyon(イネ科作物と近縁で、ゲノムサイズが小さいため遺伝学的解析に適したモデル種)に焦点を当て、この疑問に取り組む。これらの種における持続的高温ストレスへの耐性を高める順化メカニズムを探求し、イネ科植物におけるこのプロセスにおける戦略II鉄取り込みの役割を実証する。熱ストレス応答が異なる2つのB. distachyon系統の数量形質遺伝子座(QTL)解析により、熱ストレス応答の候補遺伝子としてBdTOM1を同定した。BdTOM1は高温耐性系統において高温条件下でより高発現し、2′-デオキシムギネ酸の分泌増加をもたらす。合成ムギネ酸類似体であるプロリン-2′-デオキシムギネ酸(PDMA)処理は、鉄恒常性の回復を通じて熱ストレスに伴う鉄欠乏を緩和する。対照的に、過剰な鉄処理は高温下で高レベルの活性酸素種(ROS)蓄積を引き起こす。
我々の知見は、小麦やB. distachyonのような冷涼期草本において、熱ストレス耐性を高める上で、戦略II鉄吸収機構を介した鉄恒常性の役割を明らかにした。さらに本研究は、熱誘発栄養ストレスの管理が、気候変動に対する作物の回復力を高め、地球温暖化が進む中で食糧生産を確保するための重要な戦略となり得ることを示唆している。
以下全図を訳しておいた。紙面の関係から、実際に掲載したのは図1,2,5のみである。
図1 | 高温への長期曝露後に小麦で生じる鉄欠乏症。a 水耕栽培した小麦品種「フィールダー」を、非キレート鉄[Fe2(SO4)3]を唯一の鉄源として、常温(NC、22-25℃)または高温(HC、32-35℃)条件下で2週間栽培。スケールバー:5 cm。b NCまたはHC下で2週間生育させた小麦の地上部生重量。c NCまたはHC下で2週間生育させた第2完全伸展葉の単純比色素指数(SRPI)、d 同葉の光化学反射指数(PRI)。e NCまたはHC下で2週間生育させた幼苗の第2完全伸展葉身からの最大量子収量(Fv/Fm)。f NCまたはHC下で2週間生育させた幼苗の葉身における指定金属元素の相対濃度。g 長期高温ストレスの有無による新出葉の比較。
f 2週間のNCおよびHC条件下で生育した幼苗の葉身における指定金属元素の相対濃度。g 長期高温ストレスの有無による幼苗の新生葉。幼苗は非キレート鉄を用いた水耕栽培でNCまたはHC条件下で4週間生育させた。
スケールバーは5 cm。h 4週間NCまたはHC下で生育した小麦苗の葉における鉄欠乏マーカー遺伝子TaIRO2.6の相対発現量。データは平均値±SEM(n = 4:b, e, f;n = 6:c, d;n = 3:h)。
*P < 0.05 および **P < 0.001(NCとHC間の両側Studentのt検定による)。有意差の正確なP値は図中に記載。ソースデータはソースデータファイルとして提供。
図2 | ブラキポディウム・ディスタキオンにおける高温ストレス耐性のQTLマッピング。a 非キレート鉄[Fe2(SO4)3]添加下で通常温度(NC)または高温(HC)条件下で栽培したB. distachyon系統Bd21およびBd21-3。人工フェノール樹脂ブロック(非キレート鉄 2.5mg Fe L-1 含有)で3週間培養した二葉期苗。スケールバー:5 cm。(a) の苗における相対草丈 (b) および鮮重 (c)。HC値はNC値に対する正規化値(HC/NC)。n = 6(生物学的複製)。d 4週間NCまたはHC下で生育した最初の完全展開葉における単純比色素指数(SRPI)と光化学反射指数(PRI)。n = 5(生物学的複製)。e 4週間のNCまたはHC条件下で生育した最初の完全展開葉における鉄欠乏マーカー遺伝子BdIRO2.1の相対発現量。n = 3、生物学的複製。f 4週間のNCまたはHC条件下で生育した苗の葉鉄含有量。n = 4、生物学的複製。g Bd21ゲノム内のSNP Bd4accM253とBd4accM302間の組換え近交系(RIL)の遺伝子型。SNP位置は組換え番号と共に上部に示される。オレンジ:Bd21断片;青:Bd21-3断片。h 25日間の持続高温処理後のBd21-3 × Bd21交雑RILの地上部生重量。n = 4、生物学的複製。i SNPマーカー間(Bd4accM281とBd4accM291)の32.1 kb候補領域。j 非キレート鉄下での高濃度(HC)条件で2週間水耕栽培したB. distachyon Bd21およびBd21-3幼苗における2′-デオキシムギネ酸(DMA)の分泌量および根部蓄積量(n=3、技術的反復)。データは平均値±標準誤差(SEM)を示す。*P<0.05 および **P < 0.001(両側スチューデントのt検定:b-d、g、jではBd21とBd21-3間、eではNCとHC間、hではBd21とRILs間)。有意差の正確なP値はグラフに示されている。ソースデータはソースデータファイルとして提供されている。
図3 | BdTOM1はBrachypodium distachyonにおける鉄欠乏耐性を付与する。
a 2.5mg Fe L-1の非キレート鉄[Fe2(SO4)3]を含む人工フェノール樹脂ブロック中で
高温条件下(HC)で3週間培養したBd21およびBd21-3系統における
BdTOM1およびBdTOM3の発現量。各時点において、最初に完全に展開した葉と根組織を採取した。*P < 0.05; **P < 0.001(両側Studentのt検定、Bd21対Bd21-3; n = 3、生物学的複製)。b 2.5mg Fe L-1 キレート鉄(Fe-EDTA)添加または鉄無添加(Fe-free)条件下でNCまたはHC環境下において1週間水耕栽培した幼苗におけるBdTOM1およびBdTOM3の発現量。*P<0.05; **P<0.001(両側スチューデントのt検定、Bd21対Bd21-3; n = 3、生物学的複製)。c 2.5mg Fe L-1 非キレート鉄(NC)単独、または30 μMプロリン-2′-デオキシムギネ酸(PDMA)添加(NC + PDMA)の条件下で、人工フェノール樹脂ブロック内で4週間培養したBd21、Bdtom1、Bdtom3植物。地上部生重量(d)および葉鉄含有量(e)。Bd21、Bdtom1、Bdtom3 植物を、2.5mg Fe L-1 非キレート鉄単独、または30 μMプロリン-2′-デオキシムギン酸(PDMA)添加下でNC条件下4週間培養した人工フェノール樹脂ブロック内での測定値。異なる小文字は群間の有意差を示す [一元配置分散分析; d: F (9, 29) = 16.01, P = 5.11×10-9, n =3-4; e: F (9, 30) = 15.68, P=4.41×10-9, n=4; Tukey’s HSD test, two-sided, adjusted for multiple comparisons]. f 鉄欠乏マーカーBdIRO2.1および熱ショックタンパク質BdHSP70-2の発現量。Bd21、Bdtom1、Bdtom3の幼苗を、非キレート鉄2.5mg L-1を含む人工フェノール樹脂ブロック中で4週間、非ストレス条件(NC)下で培養した際の根における発現量。*P<0.05, **P < 0.001 (両側スチューデントのt検定、Bd21対各変異体系統; n = 4, 生物学的複製)。データは平均値±SEMを示す。有意差の正確なP値はグラフ上に表示。ソースデータはソースデータファイルとして提供。
図4 | ブラキポディウム・ディスタキオン系統におけるBdTOM1多型
a 153系統のB. distachyon(NCまたはHC条件下栽培)におけるBdTOM1非同義SNP変異と幼苗生重量の関連性(補足データ1) 合計153のB. distachyon系統は、BdTOM1遺伝子型に基づき2つのハプロタイプ群、Bd21型(n = 52)とBd21-3型(n = 101)に分類された(補足データ2)。オレンジ:Bd21型;青:Bd21-3型。中央線は中央値、箱線は25%および75%四分位点を表し、ひげ線は四分位範囲の1.5倍以内の最小値と最大値を示し、バイオリン形状はデータ分布密度を示す。両群間の差異評価にはウィルコクソン順位和検定を実施した。b トルコにおけるBdTOM1ハプロタイプの地理的分布。各アクセッションの緯度経度を表示。Bd21およびBd21-3アクセッションはイラク・サラクディン近郊で採集。50のBd21型アクセス(オレンジ)および97のBd21-3型アクセス(青)のサンプリング地点は、既発表データ72(補足データ3)から取得した。画像は緯度33°~43.5°、経度25°~50°を示す。ソースデータはソースデータファイルとして提供されている。
図5 | 合成デオキシムギネ酸プロリン-2′-デオキシムギネ酸は高温ストレスに対する植物の耐性を高める。a-d B. distachyon Bd21、フェノール樹脂ブロック(非キレート鉄[Fe2(SO4)3] 2.5mg Fe L-1含有)中で、30μM プロリン-2′-デオキシムギン酸(PDMA)存在下(+)または非存在下(-)で、NCまたはHC条件下で4週間培養した植物。a 植物の代表的な画像。b 完全展開した最初の葉からの単純比色素指数(SRPI)および光化学反射指数(PRI)値。c 地上部生重量。d 葉鉄含有量。n=9, 3, 4(b-dそれぞれ)。e, f 4葉期Bd21苗を、30μM PDMA存在下または非存在下で、常温(NC)または高温(HC)条件下において2.5mg Fe L-1の非キレート鉄で1日間処理した。最初の完全展開葉と根組織を採取し、RNA-seq解析を行った。e Fe欠乏関連遺伝子発現。各行は1つの遺伝子を表し、色はlog2正規化TPM値による平均発現レベルを示す。発現レベルが低いものは黒、高いものは黄色で示される。f 光化学系およびクロロフィル代謝関連遺伝子発現。高温応答で発現が変化した遺伝子を示す。正のzスコア(オレンジ)は平均値より大きい値を示し、負のzスコア(青)は平均値より小さい値を示す。g-j 2.5mg Fe L-1の非キレート鉄を含む水耕栽培環境下で、30μM PDMA存在下(+)または非存在下(-)において、高光照条件(HC)で4週間培養した小麦‘Fielder’へのPDMA処理効果。g 植物の代表的な画像。h 最初に完全に展開した葉におけるTaIRO2.6の発現。i 最初に完全に展開した葉からのSRPIおよびPRI値。j 葉鉄含有量。n=3(h, j)、n=7(i)。データは平均値±SEM。*P<0.05、**P<0.001(PDMA有無比較の二項検定)。サンプルは生物学的に独立した反復実験。有意差の正確なP値は各グラフに表示。ソースデータはSource Dataファイルとして提供。
図6|高温条件下におけるBrachypodium distachyonへのプロリン-2′-デオキシムギネ酸および鉄毒性の用量依存的効果
a 2.5mg Fe L-1の非キレート鉄[Fe2(SO4)3]を添加した水耕栽培 B. distachyon Bd21株
[Fe2(SO4)3]を添加した水耕栽培条件下で、プロリン-2′-デオキシムギネ酸(PDMA)を
指定濃度で添加(+)または非添加(-)し、高温条件(HC)下で3週間培養した。キレート鉄(Fe-EDTA)処理では、2.5mg Fe L-1のFe-EDTAを添加した水耕栽培条件下でHC下3週間培養した。スケールバー:5cm。b 完全展開した最初の葉身から測定した単純比色素指数(SRPI)と光化学反射指数(PRI)。*P<0.05, **P < 0.001(両側t検定、PDMA処理有無の比較;n = 9、生物学的複製)。c 根における鉄欠乏マーカー遺伝子BdIRO2.1および鉄過剰マーカー遺伝子Bdフェリチンの発現。植物は2.5mg Fe L-1の非キレート鉄±PDMA(3、30、150、または300 μM)またはFe-EDTA下で高濃度(HC)で3週間培養した。*P<0.05, **P < 0.001(両側Studentのt検定、各種PDMA処理群と非処理群の比較;n = 3、生物学的複製)。d 高温処理は鉄過剰感受性を増大させる。植物をNCまたはHC条件下で、Fe-EDTAを通常濃度(45μM、2.5mg Fe L-1)または100倍高濃度(4.5mM、250mg Fe L-1)で2週間培養した。3,3′-ジアミノベンジジン染色により、若い葉においてH2O2の蓄積が検出された。スケールバー、5 cm。e 鉄キレート剤を正常濃度(×1、45 μM)または100倍高濃度(×100、4.5 mM)で1週間処理したNCまたはHC条件下で生育した最初の完全展開葉における鉄過剰マーカー遺伝子BdFerritinの発現。*P < 0.05(両側Studentのt検定、通常鉄濃度と高鉄濃度下で生育した植物を比較;n = 6、生物学的複製)。データは平均値±SEMを示す。有意差の正確なP値は各グラフに表示。ソースデータはソースデータファイルとして提供。

図1 高温への長期曝露後に小麦で生じる鉄欠乏症
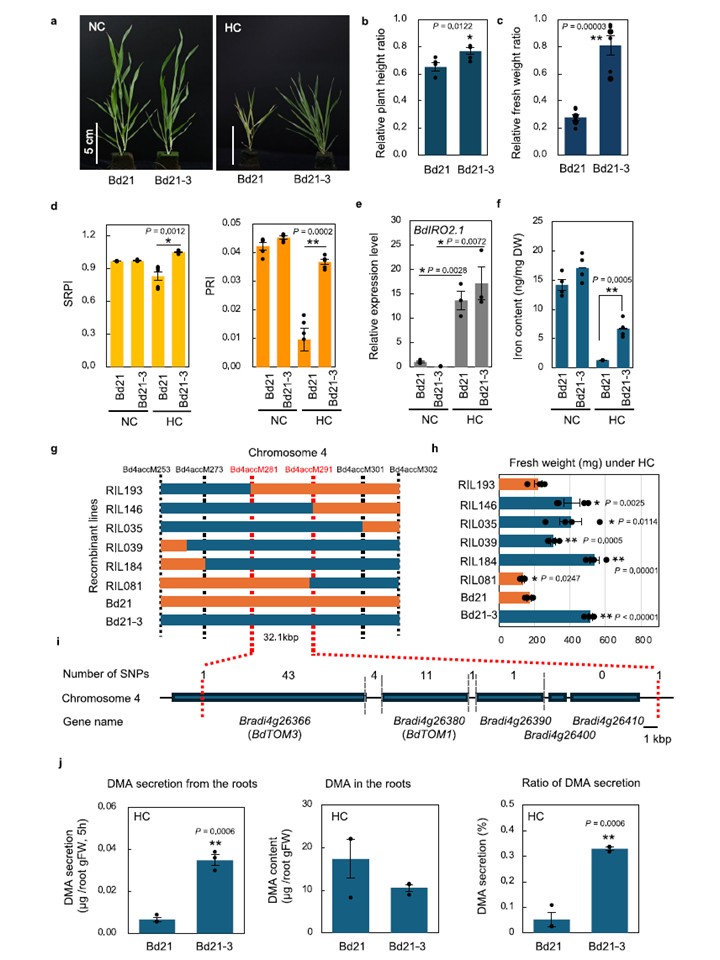
図2 B. distachyonにおける高温ストレス耐性のQTLマッピング
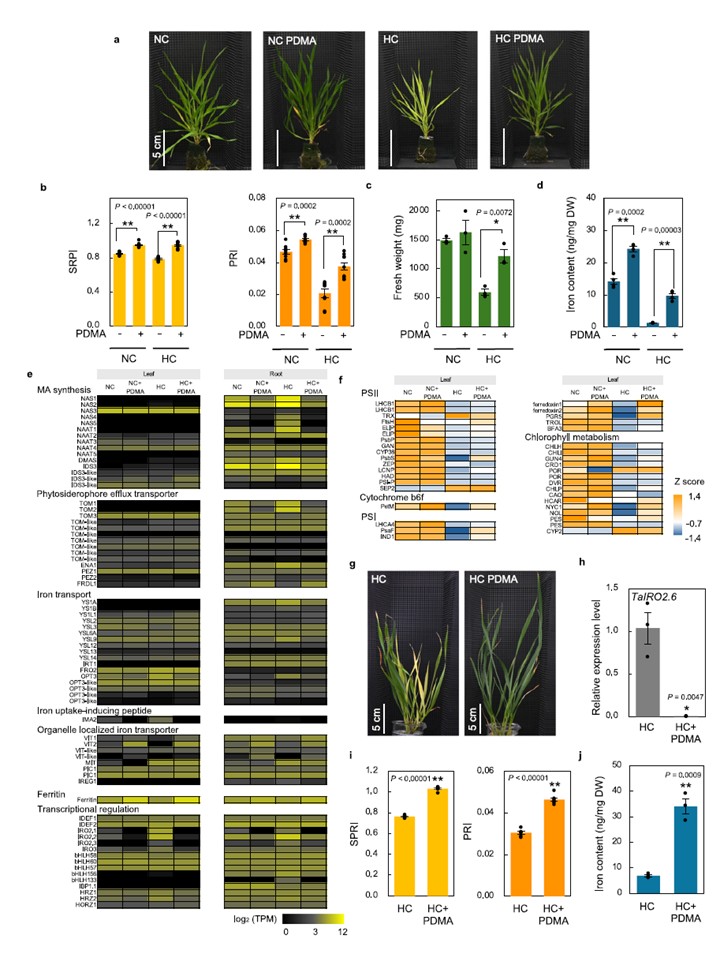
図5 合成デオキシムギネ酸プロリン-2′-デオキシムギネ酸は高温ストレスに対する植物の耐性を高める