僫僲SIMS偲摨埵懱昗幆朄傪梡偄偨怉暔揝僉儗乕僩嵻僯僐僠傾僯儈儞偺嬻娫暘晍夝愅傊偺墳梡
僫僲SIMS偲摨埵懱昗幆朄傪梡偄偨怉暔揝僉儗乕僩嵻僯僐僠傾僯儈儞偺嬻娫暘晍夝愅傊偺墳梡
Application of NanoSIMS and isotopic labelling to analyze spatial distribution of the plant iron chelator nicotianamine
Nozoye T, Takeuchi M, Hollmann F, Clemens S,
Journal of Biological Chemistry (2025), doi: https://doi.org/10.1016/j.jbc.2025.110478.
梫栺
僯僐僠傾僫儈儞乮NA乯偼丄怉暔懱偍傛傃嵶朎撪偱偺揝乮Fe乯偺桝憲偲堏摦傪巌傞揝僉儗乕僩嵻偱偡丅NA偼丄怉暔偵偍偗傞揝寚朢僔僌僫儖揱払場巕偲偟偰揝偺峆忢惈堐帩偵娭梌偟偰偄傞偲峫偊傜傟偰偄傑偡丅杮尋媶偱偼丄崅暘夝擻擇師僀僆儞幙検暘愅朄乮NanoSIMS乯傪梡偄偰丄怉暔撪偺埨掕摨埵懱昗幆NA偺暘晍傪壜帇壔偡傞偙偲偱丄NA傪夘偟偨揝偺桝憲丄堏摦丄偍傛傃揝偺峆忢惈堐帩偺儊僇僯僘儉傪夝柧偡傞偙偲傪栚揑偲偟傑偟偨丅埨掕摨埵懱昗幆NA乮15N-NA乯偼丄峺曣偵偍偗傞NA崌惉峺慺偺堎庬敪尰偵傛傝惗惉偝傟丄栰惗宆乮WT乯僩儅僩怉暔偲NA寚懝曄堎懱chloronerva乮chln乯偵嫙媼偝傟傑偟偨丅NanoSIMS偺寢壥丄15N-NA偺僔僌僫儖偼丄WT偲chln偺椉曽偱揝偑拁愊偡傞摫娗廃曈偺慻怐偵廤拞偟偰偄傑偟偨丅摉尋媶偺寢壥偼丄NanoSIMS偑怉暔懱撪偱偺NA嬊嵼壔傪捈愙挷嵏偡傞嫮椡側僣乕儖偱偁傞偙偲傪帵偟偰偄傑偡丅
埲壓榑暥偺娞偺晹暘偩偗栿偟傑偟偨丅
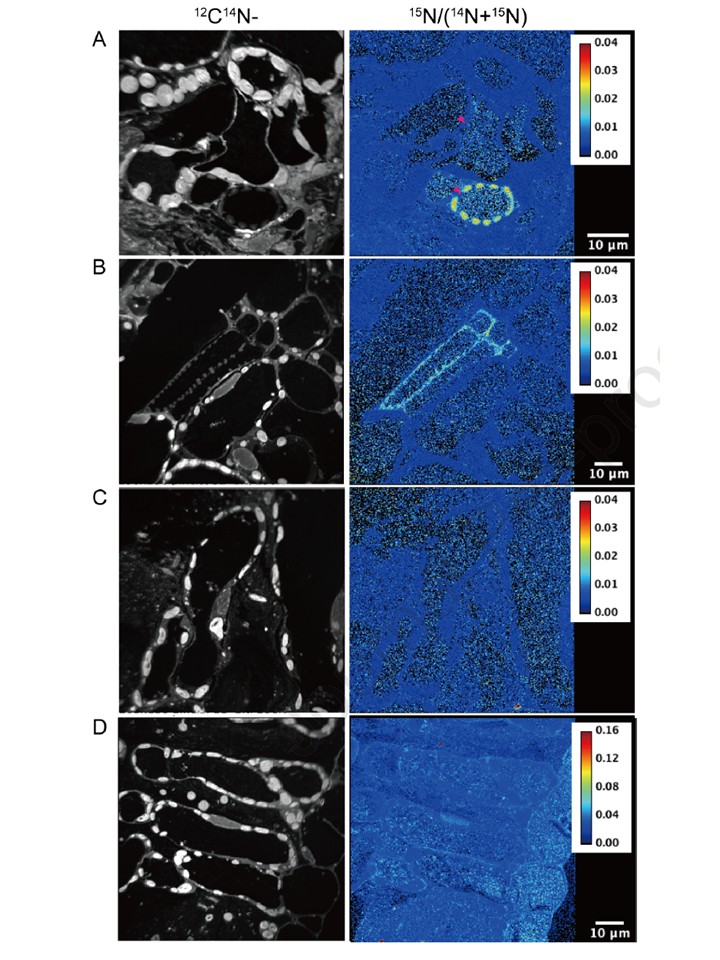
僫僲SIMS夝愅偵傛傝丄15N-NA偑栘晹廃曈偺嵶朎偵拁愊偟偰偄傞偙偲偑敾柧偟丄揝偺嬊嵼壔偲堦抳偡傞寢壥偱偟偨丅15N壔崌暔偑宻偺婎晹偵嫙媼偝傟偨偨傔丄栘晹廃曈偺嵶朎偵偍偗傞15N偺崅拁愊偼丄15211 N-NA偑栘晹偐傜媧廂偝傟丄偦偺屻廃曈偺嵶朎傊桝憲偝傟偨壜擻惈傪帵嵈偟偰偄傑偡乮恾2乯丅崻偺嵶朎暻偵偍偗傞Fe擹搙偼丄chln偱偼WT偵斾傋偰嬌傔偰掅偔丄嵶朎幙偲塼朎偵偍偗傞Fe擹搙偼chln偱WT傛傝傕崅偐偭偨偙偲偑曬崘偝傟偰偄傑偡丅NA傪揔梡偡傞偲丄chln偲WT偺娫偺偙傟傜偺嵎偑徚幐偟丄NA偑傾億僾儔僗僩偺Fe偺梟夝偲棙梡偵昁梫偱偁傞偙偲傪帵嵈偟偰偄傑偡丅X慄媧廂嬤曈峔憿乮XANES乯傕丄chln偺梩柆偵偍偗傞擇壙揝偺懚嵼検偑WT傗梩柆娫椞堟傛傝傕掅偄偙偲傪帵偟丄NA偑僩儅僩梩偺梩柆偵偍偗傞揝偺擇壙宍懺偺堐帩偵娭梌偟偰偄傞壜擻惈傪帵嵈偟偰偄傑偡丅摨條偵丄揝偺嬊嵼偼庡偵崻丄宻丄梩偺寣娗宯偵娤嶡偝傟傑偟偨乮恾3-5乯丅嵶朎撪偱偼丄塼朎撪偺15N僔僌僫儖偼嵶朎幙傛傝傕崅偄孹岦偵偁傝丄15N-NA偑嵶朎偵庢傝崬傑傟丄偦偺屻塼朎傊桝憲偝傟傞壜擻惈傪帵嵈偟偰偄傑偡乮恾2乯丅
幚嵺丄峈NA峈懱傪梡偄偨柶塽愼怓偵傛傝丄NA偼揝偑廫暘側忦審壓偍傛傃揝偑晄懌偡傞忦審壓偱嵶朎幙偵嬊嵼偟丄
揝偑朙晉側忦審壓偱偼彫朎懱偵堏摦偡傞偙偲偑帵嵈偝傟傑偟偨丅NA偑揝偺
僗僇儀儞僕儍乕偲偟偰婡擻偟丄塼朎傊偺晻偠崬傔偵傛傝嵶朎傪曐岇偡傞壜擻惈偑採彞偝傟傑偟偨丅偟偐偟丄15N-NA傪娷傓僒儞僾儖偲丄15N偱攟梴偝傟偨VC偺拪弌暔傪娷傓僒儞僾儖偺椉曽偱丄嵶朎幙偲塼朎偵偍偗傞15N僔僌僫儖偑娤嶡偝傟傑偟偨乮恾2乯丅摨條偵丄WT偲chln偺椉曽偺僒儞僾儖偵偍偄偰丄15N偺擹搙偼僒儞僾儖娫偱嵎偑偁傝傑偣傫偱偟偨丅偝傜偵丄娤嶡偝傟偨僔僌僫儖偼丄枹擣幆偺NA戙幱暔偵傛偭偰惗惉偝傟偨壜擻惈傕偁傝傑偡丅偙傟偵懳墳偡傞偨傔丄15N傪摨埵懱僩儗乕僒乕偲偟偰巊梡偡傞儊僞儃儘儈僋僗傾僾儘乕僠偑昁梫偲側傝傑偡丅暋悢偺慻怐偵偍偄偰暋悢偺忦審壓偱nanoSIMS傪揔墳偝偣傞偙偲偱丄NA偺揝桝憲偵偍偗傞栶妱傪柧傜偐偵偡傞壜擻惈偑偁傝傑偡丅
寢榑偲偟偰丄杮尋媶偼丄NanoSIMS偲摨埵懱昗幆偺慻傒崌傢偣偑丄嬥懏僉儗乕僩嵻NA偺傛偆側暘巕偺嵶朎撪偍傛傃慻怐儗儀儖偱偺堏摦傪摿掕偡傞戝偒側壜擻惈傪桳偡傞偙偲傪帵偟傑偟偨丅偝傜側傞夵慞偼丄NA偺惗棟妛揑棟夝傪怺傔傞偺偵峷專偡傞壜擻惈偑偁傝傑偡丅
恾偺杴椺乮恾偼1丆2丆3偟偐帵偣側偄偑丄係丆俆傕栿偟偰偍偄偨乯
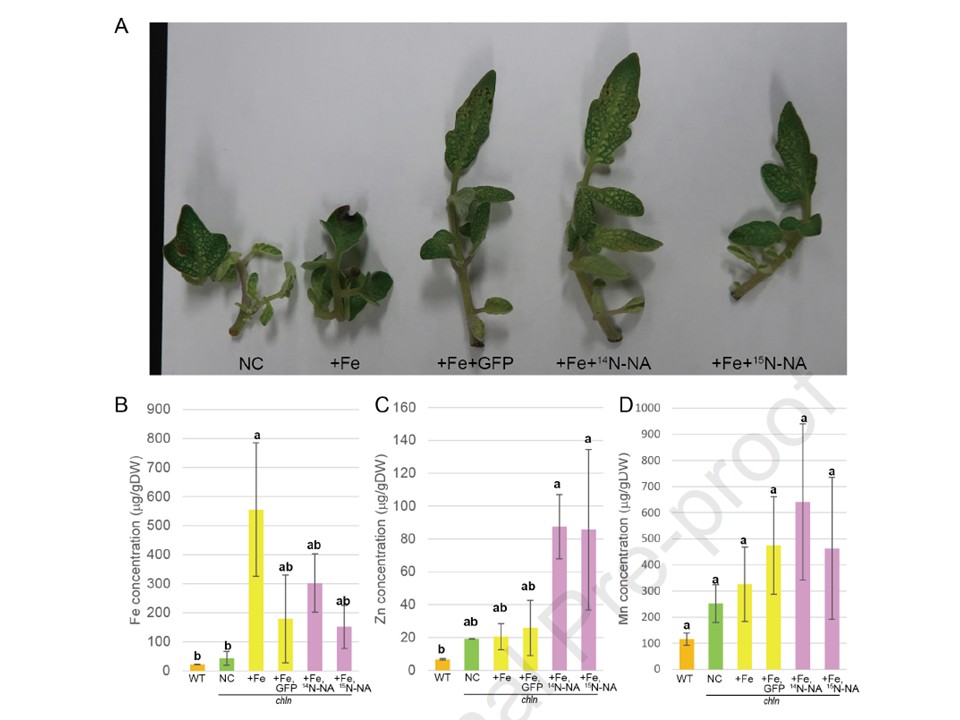
恾1. 峺曣拪弌暔傪梡偄偨僩儗乕僒乕幚尡丅僋儘儘僱儖僶乮chln乯偺宻偼悈峩嵧攟偱攟梴偝傟丄僩儗乕僒乕幚尡偵梡偄傜傟偨乮A乯丅宻偼丄揝晄娷偺悈峩攟梴塼乮堿惈懳徠丟NC乯丄揝娷桳乮+Fe乯偍傛傃GFP峺曣拪弌暔乮+Fe+GFP乯丄揝偲GFP-AtNAS2峺曣拪弌暔乮14N-NA娷桳乯傪娷傓梟塼乮+Fe+14N-NA乯偵怹捫偝傟偨丅乮+Fe+14 N-NA乯丄傑偨偼Fe偲GFP-AtNAS2峺曣拪弌暔乮15N-NA娷桳丟+Fe+15NNA乯偵怹捫偟偨丅2擔屻丄宻偺Fe乮B乯丄Zn乮C乯丄Mn乮D乯擹搙傪暘愅偟偨丅悈峩嵧攟偱攟梴偝傟偨栰惗宆乮WT乯偺宻偺嬥懏擹搙傕應掕偟偨丅 抣偼3偮偺惗暔妛揑暋惢暯嬒傪昞偡丅僄儔乕僶乕偼昗弨曃嵎傪帵偡丅DW偼姡憞廳検丅堎側傞暥帤偼Tukey-Kramer HSD専掕乮n = 3丄P < 0.1乯偵傛傞桳堄嵎傪帵偡丅
恾2. 崅夝憸搙擇師僀僆儞幙検暘愅乮NanoSIMS乯夋憸偱帵偝傟傞僩儅僩怉暔偺梩乮栰惗宆丟A丄C丄僋儘儘僱儖僶丟B丄D乯偵偍偗傞15N偺暘晍丅僩儗乕僒乕幚尡偵偍偄偰丄15N-NA傪娷傓GFP-AtNAS2峺曣偺拪弌暔乮A丄B乯偍傛傃15N傪娷傓攟抧偱攟梴偝傟偨VC峺曣偺拪弌暔乮C丄D乯傪巊梡偟丄15N偺暘晍傪暘愅偟偨丅僶乕丄10 兪m丅
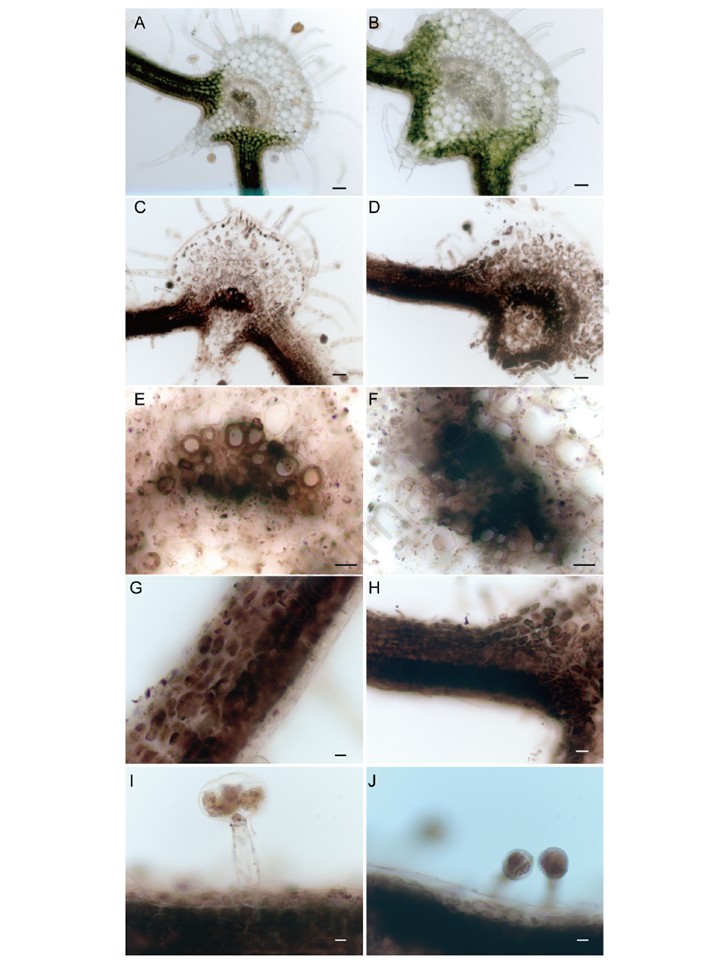
恾3. 栰惗宆乮WT丟C丄E丄G丄I乯偲僋儘儘僱儖僶乮chln丟D丄F丄H丄J乯偺梩偵偍偗傞Fe偺嬊嵼壔傪丄Perl偺僾儖僔傾儞僽儖乕-DAB朄偱帵偟偨丅
張棟慜偺WT乮A乯傑偨偼chln乮B乯偺梩偺墶抐柺傕偦傟偧傟帵偝傟偰偄傞丅戝偒側寣娗懇乮E丄F乯丄 拞梩嵶朎乮G丄H乯丄偍傛傃栄忬懱乮I丄J乯偑奼戝昞帵偝傟偰偄傞丅僗働乕儖僶乕丄50 兪m乮A-D乯傑偨偼20 兪m乮E-J乯丅
恾4. 栰惗宆乮WT丟C丄E丄G丄I丄K乯偲僋儘儘僱儖僶乮chln丟D丄F丄H丄J丄L乯偺宻偵偍偗傞Fe偺嬊嵼壔傪丄Perl偺僾儖僔傾儞僽儖乕-DAB朄偱帵偟偨丅WT乮A乯傑偨偼chln乮B乯偺張棟慜偺宻偺抐柺傕偦傟偧傟帵偝傟偨丅戝偒側寣娗懇乮E, F乯丄昞旂乮G, H乯丄旂憌乮I, J乯丄偍傛傃栄忬懱乮K, L乯偑奼戝昞帵偝傟偰偄傞丅僗働乕儖僶乕丗50 兪m乮A-D乯傑偨偼20 兪m乮E-L乯丅
恾5. 栰惗宆乮WT丟C丄E丄G丄I乯偲僋儘儘僱儖僶乮chln丟D丄F丄H丄J乯偺崻偵偍偗傞Fe偺嬊嵼壔傪Perl偺僾儖僔傾儞僽儖乕-DAB朄偱帵偟偨丅張棟慜偺崻偺廲抐柺乮WT丗A丄chln丗B乯傕偦傟偧傟帵偝傟偰偄傞丅Fe偺嬊嵼壔偼丄廲抐柺乮C丄D乯偲墶抐柺乮E丄F乯偱娤嶡偝傟偨丅寣娗懇乮G丄H乯偲旂憌乮I丄J乯偼奼戝昞帵偝傟偰偄傞丅僗働乕儖僶乕丗1 mm乮A丄B乯傑偨偼20 兪m乮C-J乯丅
恾侾
恾俀
恾俁