RNA僿儕僇乕僛偼揝僔僌僫儖惂屼場巕偲嫤挷偟偰僔儘僀僰僫僘僫偺姦椻僗僩儗僗傪娚榓偡傞
偙偺榑暥偼彫惗偺抦傞偲偙傠偱偼姦椻僗僩儗僗偲揝偲偺娭學傪柧傜偐偵偟偨嵟弶偺榑暥偱偼側偄偩傠偆偐丅姦椻僗僩儗僗偺暘巕惗暔妛揑側暋嶨側抦幆偑側偄偨傔偵丄巆擮側偑傜彫惗偵偼偙偺榑暥偼枹徚壔偺傑傑偱偁傞丅DEAD-box RNA僿儕僇乕僛偱偁傞RH24傪敪尒偟偨偙偲偵怴婯惈偑偁傞傜偟偄丅
RNA僿儕僇乕僛偼揝僔僌僫儖惂屼場巕偲嫤挷偟偰僔儘僀僰僫僘僫偺姦椻僗僩儗僗傪娚榓偡傞
An丂RNA丂helicase coordinates with iron signal regulators to alleviate chilling stress in Arabidopsis
Yingying Xing, Yawen Li, Xinmeng Gui, Xianyu Zhang, Qian Hu, Qiqi Zhao, Yongli Qiao, Ning Xu & Jun Liu
Nature Communications volume 16, Article number: 3988 (2025)
梫巪
姦椻僗僩儗僗偼丄怉暔偺敪堢偲惉挿傪梷惂偡傞庡梫側娐嫬僗僩儗僗偺傂偲偮偱偁傞丅変乆偺埲慜偺尋媶偱偼丄僔儘僀僰僫僘僫 怉暔偵偍偄偰丄愽嵼揑側揝僙儞僒乕BTS乮BRUTUS乯偑壏搙墳摎偵娭梌偟偰偄傞偙偲偑帵偝傟偨丅偟偐偟丄怉暔偺揝儂儊僆僗僞僔僗偑壏搙曄摦偵懳偡傞怉暔偺墳摎偵娭梌偟偰偄傞偐偳偆偐偼晄柧偱偁傞丅杮尋媶偱偼丄BTS曄堎懱bts-2偑掅壏僗僩儗僗偵姶庴惈偑偁傝丄偦偺姶庴惈偼揝偺拁愊偵婲場偡傞偙偲傪敪尒偟偨丅bts-2偺僒僾儗僢僒乕僗僋儕乕僯儞僌偺寢壥丄bts-2偺掅壏姶庴惈傪姰慡偵梷惂偡傞DEAD-box RNA僿儕僇乕僛偱偁傞RH24傪敪尒偟偨丅RH24偼掅壏壓偱拁愊偟丄揝 惂屼場巕ILR3乮IAA-儘僀僔儞懴惈3乯mRNA傪姫偒栠偟丄ILR3僞儞僷僋幙 儗儀儖傪憹壛偝偣傞丅嫽枴怺偄偙偲偵丄RH24偼ILR3傪憡暘棧偟偨嬅弅懱拞偵晻偠崬傔丄ILR3偑夘嵼偡傞揝夁忚晧壸傪寉尭偡傞丅偝傜偵BTS傑偨偼掅壏張棟偵傛傝丄嬅弅懱偺宍惉偑懀恑偝傟偨丅偟偨偑偭偰丄RH24偲BTS偼嫤挷揑偵ILR3傪惂屼偟丄椻壏僗僩儗僗壓偱偺揝偺庢傝崬傒傪梷惂偡傞丅変乆偺敪尒偼丄RNA僿儕僇乕僛RH24偲BTS偑丄壏搙曄摦偵懳墳偟偰怉暔偺揝 儂儊僆僗僞僔僗傪堐帩偡傞偨傔偵丄ILR3傪旝挷惍偟偰偄傞偙偲傪柧傜偐偵偟偨丅
偼偠傔偵
姦椻僗僩儗僗偼庡梫側娐嫬僗僩儗僗偺堦偮偲偟偰丄怉暔偺惉挿丄敪堢丄抧棟揑暘晍傪梷惂偡傞丅抧媴婯柾偺婥岓曄摦偵傛傝丄嬌抂側掅壏尰徾偑昿斏偵敪惗偟丄擾嬈偵敎戝側宱嵪揑懝幐傪傕偨傜偟偰偄傞丅掅壏姶庴惈偺怉暔偵偲偭偰丄掅壏僗僩儗僗偼捠忢丄惗挿偍傛傃惗怋偺椉抜奒偵偍偗傞惉挿偲敪払傪慾奞偟丄偟偽偟偽惉挿偺慾奞丄敪夎偺掅壓丄梩偺僋儘儘僔僗丄偝傜偵偼怉暔偺嵟廔揑側屚巰傪敽偆丅掅壏壓偱惗偒巆傞偨傔偵丄怉暔偼掅壏偵傛傞僗僩儗僗傪夵慞偡傞暋嶨側挷愡婡峔傪恑壔偝偣偰偒偨丅
掅壏僗僩儗僗偼丄嵶朎枌偺棳摦惈傪掅壓偝偣丄巁壔娨尦忬懺傪曄壔偝偣丄岝崌惉傗峺慺妶惈傪慾奞偟丄偟偽偟偽妶惈巁慺庬偺拁愊傗揹夝幙楻弌偺忋徃傪堷偒婲偙偡偙偲偑抦傜傟偰偄傞丅C-儕僺乕僩寢崌場巕乮CBFs乯堚揱巕側偳偺戙昞揑側揮幨場巕偼丄掅壏忦審壓偱媫懍偵桿摫偝傟偨丅偙傟傜偺桿摫偝傟偨CBFs偼丄堦楢偺壓棳偺掅壏惂屼乮COR乯堚揱巕傪捈愙妶惈壔偟丄寢壥偲偟偰怉暔偺懴椻惈傪崅傔傞怉暔偺懴姦惈偵偍偄偰CBF偑廳梫側栶妱傪壥偨偟偰偄傞偵傕偐偐傢傜偢丄CBF偵傛偭偰惂屼偝傟偰偄傞COR堚揱巕偼慡懱偺栺10%偵偡偓側偄丅幚嵺丄崅師偺cbfs曄堎懱偱偝偊丄埶慠偲偟偰偐側傝偺掅壏懴惈傪桳偟偰偍傝丄CBF偵埶懚偟側偄僔僌僫儖揱払宱楬偑怉暔偺掅壏懴惈偺惂屼偵娭梌偟偰偄傞偙偲偑帵嵈偝傟偰偄傞丅
揮幨場巕偵壛偊偰丄mRNA偺揮幨屻惂屼傕怉暔偺姦椻墳摎偵娭梌偟偰偄傞丅RNA僿儕僇乕僛偼丄mRNA偺揮幨屻惂屼傪巌傞廳梫側僞儞僷僋幙僼傽儈儕乕偺堦偮偱偁傞丅RNA僿儕僇乕僛偼丄RNA姫偒栠偟妶惈偲RNA愜傝忯傒夝彍妶惈傪夘偟偰丄惉弉RNA傪惓偟偄峔憿偵宍惉偡傞偙偲傪妋幚偵偡傞峺慺偱偁傞丅DEAD乮Asp-Glu-Ala-Asp乯儃僢僋僗RNA僿儕僇乕僛偼丄RNA僿儕僇乕僛僼傽儈儕乕偺拞偱嵟傕戝偒側僋儔僗偱偁傞丅僿儕僇乕僛僐傾僪儊僀儞偵偼9偮偺曐懚偝傟偨儌僠乕僼偑偁傞丅僿儕僇乕僛僐傾僪儊僀儞偼RNA偲ATP偺寢崌偵昁梫偱偁傝丄mRNA傪壛悈暘夝偡傞偺偵昁恵偱偁傞丅偝傜偵丄DEAD-box RNA僿儕僇乕僛偼N枛抂偲C枛抂偵壜曄挿攝楍傪帩偪丄婎幙寢崌摿堎惈傪婯掕偟偰偄傞偲採彞偝傟偰偄傞丅偙偺傛偆偵丄偦傟偧傟偺DEAD-box RNA僿儕僇乕僛偼丄mRNA偺僗僾儔僀僔儞僌丄桝憲丄東栿丄RNA僾儘僙僔儞僌側偳丄RNA戙幱偵偍偄偰儐僯乕僋側栶妱傪壥偨偟偰偄傞丅
懡偔偺DEAD-box RNA僿儕僇乕僛偑丄怉暔偺姦椻墳摎偵偍偄偰CBF偵埶懚偟側偄宱楬偱摥偔偙偲偑曬崘偝傟偰偄傞丅椺偊偽丄RH42/RCF1偼僾儗mRNA偺揔愗側僗僾儔僀僔儞僌傪堐帩偟丄怉暔偺懴姦惈傪惂屼偟偰偄傞丅RH25偼姦椻僗僩儗僗偵傛偭偰桿摫偝傟丄RH25偺夁忚敪尰偼怉暔偺搥寢懴惈傪崅傔傞丅RH38偼mRNA偺桝憲偵廳梫側栶妱傪壥偨偟丄怉暔偺姦椻僗僩儗僗墳摎偵娭梌偟偰偄傞丅RH7/PRH75偼怉暔偑姦椻僗僩儗僗偵偝傜偝傟傞偲18S pre-rRNA偺僾儘僙僔儞僌偵娭梌偡傞丅偙傟傜偺徹嫆偼丄DEAD-box RNA僿儕僇乕僛偑掅壏僗僩儗僗傊偺揔墳偵偍偄偰懡條偵怉暔傪彆偗偰偄傞偙偲傪帵偟偰偄傞丅
揝乮Fe乯偼丄傎偲傫偳偡傋偰偺惗暔偵偲偭偰嵟傕廳梫側旝検尦慺偱偁傞丅僋儘儘僼傿儖惗崌惉丄屇媧丄DNA暋惢丄僄僱儖僊乕惗嶻側偳丄懡條側嵶朎僾儘僙僗偵偍偗傞懡偔偺廳梫側峺慺偺曗峺慺偱偁傞丅揝偺寚朢偼丄僸僩偱偼昻寣傪丄怉暔偱偼僋儘儘僔僗傪堷偒婲偙偡偑丄揝偺夁忚愛庢偼丄僸僪儘僉僔儖儔僕僇儖傪惗惉偟偰DNA傪懝彎偟丄僞儞僷僋幙傗帀幙傪夁巁壔偡傞僼僃儞僩儞斀墳傪桿敪偡傞偨傔丄桳奞偱偁傞丅揝偼廳梫偱偁傞偨傔丄娐嫬僗僩儗僗壓偱揝偺儂儊僆僗僞僔僗傪堐帩偡傞偙偲偼丄怉暔偺惗懚偵偲偭偰廳梫偱偁傞丅姦椻丄揝丄墫僗僩儗僗壓偱偺僀僱偺斾妑僩儔儞僗僋儕僾僩乕儉夝愅偵傛傝丄468偺嵎師揑敪尰堚揱巕乮DEG乯偑姦椻僗僩儗僗偲揝僗僩儗僗偺椉曽偱尒偮偐偭偨丅FRO2乮揝娨尦僆僉僔僟乕僛2乯偼丄揝偺庢傝崬傒偺偨傔偵揝傪Fe3+偐傜Fe2+偵娨尦偡傞揝僉儗乕僩娨尦峺慺偱偁傝丄僔儘僀僰僫僘僫偺崻偵偍偄偰僌儕僔儞儀僞僀儞偑桿摫偡傞姦椻懴惈偵昁梫偱偁傞丅僞僶僐偱偼丄梩椢懱偵僼僃儕僠儞偑拁愊偡傞偙偲偱丄掅壏偵傛傞岝慾奞偐傜怉暔傪庣偭偰偄傞丅僔儘僀僰僫僘僫偺崻偵偍偗傞揝偺拁愊憹壛偼丄儕儞巁乮Pi乯婹夓偵敽偆懴椻惈偵傕昁梫偱偁傞丅掅壏偼怉暔偺岝崌惉岠棪傪掅壓偝偣丄岝壔妛宯I乮PSI乯偵僟儊乕僕傪梌偊傞丅PSI僔僗僥儉偵偼揝偑娷傑傟偰偍傝丄掅壏僗僩儗僗偼PSI婡擻偵塭嬁傪梌偊傞偙偲偱丄揝偺峆忢惈偵塭嬁傪梌偊傞壜擻惈偑偁傞丅偙傟傜偺徹嫆偼丄揝偺儂儊僆僗僞僔僗偑怉暔偺懴椻惈偵戝偒偔娭學偟偰偄傞偙偲傪帵嵈偟偰偄傞丅偲偼偄偊丄怉暔偑偳偺傛偆偵揝儂儊僆僗僞僔僗傪挷愡偟偰懴椻惈傪崅傔偰偄傞偺偐偼丄埶慠偲偟偰晄柧偱偁傞丅
愽嵼揑側揝僙儞僒乕僞儞僷僋幙偱偁傞BRUTUS乮BTS乯偼丄揝儂儊僆僗僞僔僗偺廳梫側挷愡場巕偱偁傞丅偙偺僞儞僷僋幙偼丄僒僽僌儖乕僾偺墫婎惈僿儕僢僋僗-儖乕僾僿儕僢僋僗乮bHLH乯揮幨場巕ILR3乮IAA-儘僀僔儞懴惈3乯偲bHLH11538偺暘夝傪懀恑偡傞偙偲偵傛偭偰丄揝寚朢墳摎傪惂屼偟偰偄傞丅揝寚朢壓偱偼ILR3偑妶惈壔偝傟丄怉暔偵偍偗傞揝偺庢傝崬傒偲挋憼傪懀恑偡傞丅偟偐偟丄BTS偼ILR3傪昗揑偲偟偰暘夝偟丄怉暔偑夁忚側揝傪妉摼偡傞偺傪杊偖丅
傢傟傢傟偺埲慜偺尋媶偱丄bts-2偼壏搙偵晀姶偱偁傞偙偲偑帵偝傟偨丅杮尋媶偱偼丄BTS曄堎懱bts-2偑側偤掅壏偵晀姶側偺偐偲偄偆媈栤傪夝寛偟丄怉暔偺揝夁忚愛庢偑懴姦惈傪掅壓偝偣傞偙偲傪尒弌偟偨丅変乆偼丄bts-2偺掅壏姶庴惈偲揝僗僩儗僗懴惈偺昞尰宆傪姰慡偵梷惂偱偒傞DEAD-box RNA僿儕僇乕僛丄RH24傪敪尒偟偨丅RH24偼椻壏僗僩儗僗壓偱拁愊偡傞丅RH24偼ILR3 mRNA傪姫偒栠偟丄ILR3僞儞僷僋幙儗儀儖傪憹壛偝偣傞丅偟偐偟丄RH24偲ILR3偼椻壏僗僩儗僗壓偱憡暘棧偟偨嬅弅懱傪宍惉偟丄BTS偼嬅弅懱偺宍惉傪懀恑偡傞丅偦偺寢壥丄怉暔偼揝偺庢傝崬傒傪尭彮偝偣傞丅変乆偼丄RH24偲BTS偑ILR3傪夘偟偰揝偺儂儊僆僗僞僔僗傪惓妋偵挷愡偟丄僔儘僀僰僫僘僫偺椻壏僗僩儗僗傪娚榓偡傞偙偲傪採彞偡傞丅
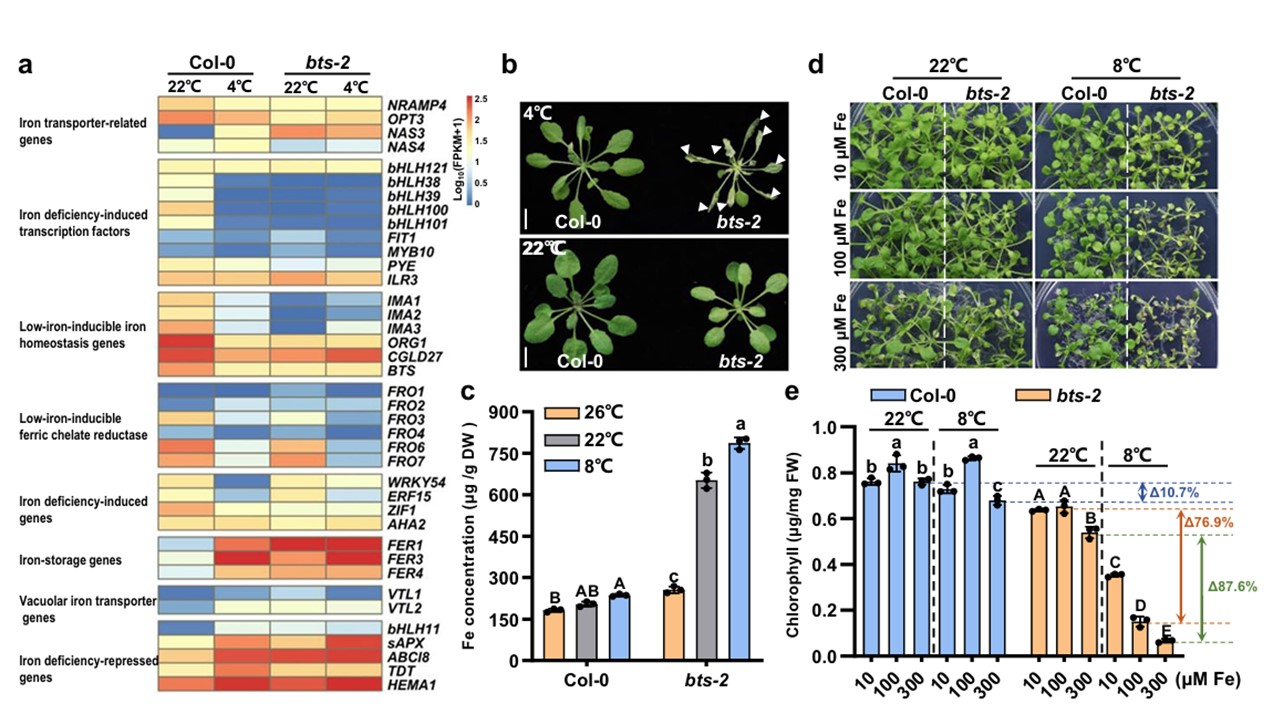
恾1丂 bts-2偺掅壏姶庴惈偼揝偺拁愊偲娭楢偟偰偄傞丅
a 4亷壓偵偍偗傞怉暔偺揝戙幱宱楬偺敪尰嵎堚揱巕乮DEG乯丅col-0偍傛傃bts-2怉暔傪22亷偱4廡娫惗堢偝偣偨屻丄4亷偱24帪娫張棟偟偨丅4廡楊偺怉暔偺梩傪僩儔儞僗僋儕僾僩乕儉傾僢僙僀梡偵僒儞僾儕儞僌偟偨丅幚尡偼2夞峴偭偨丅
b 掅壏僗僩儗僗壓偱偺bts-2怉暔偺昞尰宆丅col-0偍傛傃bts-2怉暔傪26亷偱3廡娫惗堢偝偣丄偝傜偵22亷傑偨偼4亷偱7擔娫惗堢偝偣偨丅栴報偼屚傟偨梩傪帵偡丅怉暔偼22亷偺搚忞偱2廡娫惗堢偝偣偨屻丄26亷丄22亷丄8亷偵堏偟偰偝傜偵2廡娫惗堢偝偣偨丅
們 揝擹搙偼姡憞廳検儀乕僗偱偁傝丄4廡楊偺怉暔偺梩傪僒儞僾儕儞僌偟偰揝尦慺暘愅傪峴偭偨丅奺斀暅幚尡偵偼栺20姅傪梡偄偨丅
d 揝偺夁忚嫙媼偼bts-2偺掅壏姶庴惈傪憹壛偝偣偨丅col-0偍傛傃bts-2怉暔傪22亷偺1/2MS偱2廡娫惗堢偝偣偨屻丄10, 100, 偍傛傃300兪M偺Fe(II)EDTA傪揧壛偟偨Fe寚朢1/2MS攟抧乮-Fe乯偵堏怉偟丄偦傟偧傟22亷偱1廡娫丄8亷偱2廡娫惗堢偝偣偨丅10,100,300兪M偺Fe偼丄偦傟偧傟Fe寚朢丄Fe廩懌丄Fe夁忚偺忦審偵懳墳偡傞丅
e (d)偺Col-0偍傛傃bts-2昪偺僋儘儘僼傿儖擹搙丅僋儘儘僼傿儖擹搙偺應掕偵偼丄奺斀暅幚尡偵偮偒6姅傪嵦庢偟偰梡偄偨丅
僨儖僞抣偼丄8亷偵偍偗傞僋儘儘僼傿儖擹搙偺尭彮棪傪丄22亷偵偍偗傞揝夁忚乮300兪MFe乯忦審壓偲揝夁忚乮100兪MFe乯忦審壓偱斾妑偟偨傕偺偱偁傞丅
丂丂
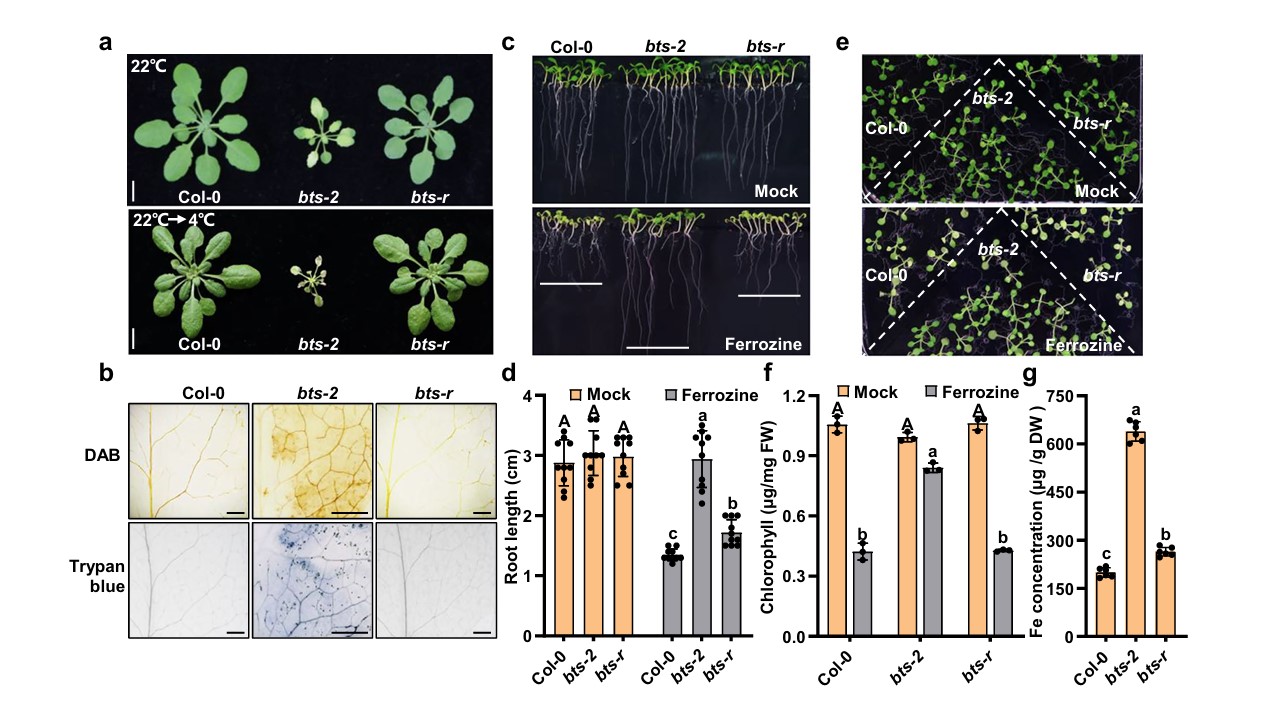
恾2|bts-r乮bts-2偺僒僾儗僢僒乕乯偼丄掅壏偱偺惗堢擻椡偍傛傃寚懝懴惈偑掅壓偟偨丅
a bts-r乮bts-2偺僒僾儗僢僒乕乯偼掅壏懴惈傪掅壓偝偣偨丅Col-0丄bts-2偍傛傃bts-r偼22亷偱4廡娫惗堢偝偣偨乮忋恾乯丅
b bbts-rd偼掅壏壓偱H2O2傪拁愊偟側偐偭偨丅22亷偱4廡娫惗堢偝偣丄偝傜偵2擔娫4亷偵堏偟偨丅梩偼DAB偲僩儕僷儞僽儖乕偱愼怓偟偨丅
c bts-rw偼晄姰慡惈偵晀姶偱偁傞丅偙傟傜偺曄堎懱偼丄100兪M偺Fe(II)-EDTA傪娷傓1/2MS攟抧偱惗堢偝偣偨丅300兪M偺僼僃儘僕儞傪1/2MS攟抧偵7擔娫 搳梌偟丄揝傪僉儗乕僩偟偨乮壓恾乯丅
d (c)偺怉暔偺崻偺挿偝偺摑寁暘愅丅
e 揝寚朢壓偵偍偗傞bts-怉暔偺昞尰宆丅 1/2MS傪22亱C 偱7擔娫惗堢偝偣丄100兪M Fe(II)-EDTA 傑偨偼300兪M僼僃儘僕儞(揝寚朢)傪娷傓1/2MS攟抧偵偦傟偧傟5擔娫堏偟偨丅
f 僋儘儘僼傿儖擹搙(e)...1姅摉偨傝6姅傪嵦庢偟丄僋儘儘僼傿儖擹搙應掕偵梡偄偨丅
Col-0丄bts-2丄bts-r偺奺僾儔儞僩偵偍偗傞揝擹搙丅 揝擹搙偺應掕偵偼丄22亷偺搚忞偱4廡娫惗堢偝偣偨怉暔偺梩傪梡偄偨丅
恾俈.f嶌嬈儌僨儖丗RH24偼ILR3偺mRNA傪姫偒栠偟丄ILR3偺拁愊傪懀恑偡傞丅姦椻僗僩儗僗壓偱偼丄RH24偑拁愊偝傟丄妀撪偵RH24-ILR3嬅弅懱傪宍惉偡傞偙偲偱ILR3傪憡暘棧懱偵晻偠崬傔傞丅BTS偲姦椻僗僩儗僗偼RH24-ILR3嬅弅懱偺宍惉傪懀恑偡傞丅偦偺寢壥丄ILR3偑惂屼偡傞揝偺庢傝崬傒偼姦椻僗僩儗僗壓偵偁傝丄偦偺寢壥丄揝夁晧壸偵傛傞懝彎偑尭庛偡傞丅昗弨揑側惗堢忦審壓偱偼丄BTS偼梀棧ILR3傪旝挷惍偟丄揝儂儊僆僗僞僔僗傪堐帩偡傞丅
恾1.
恾2.
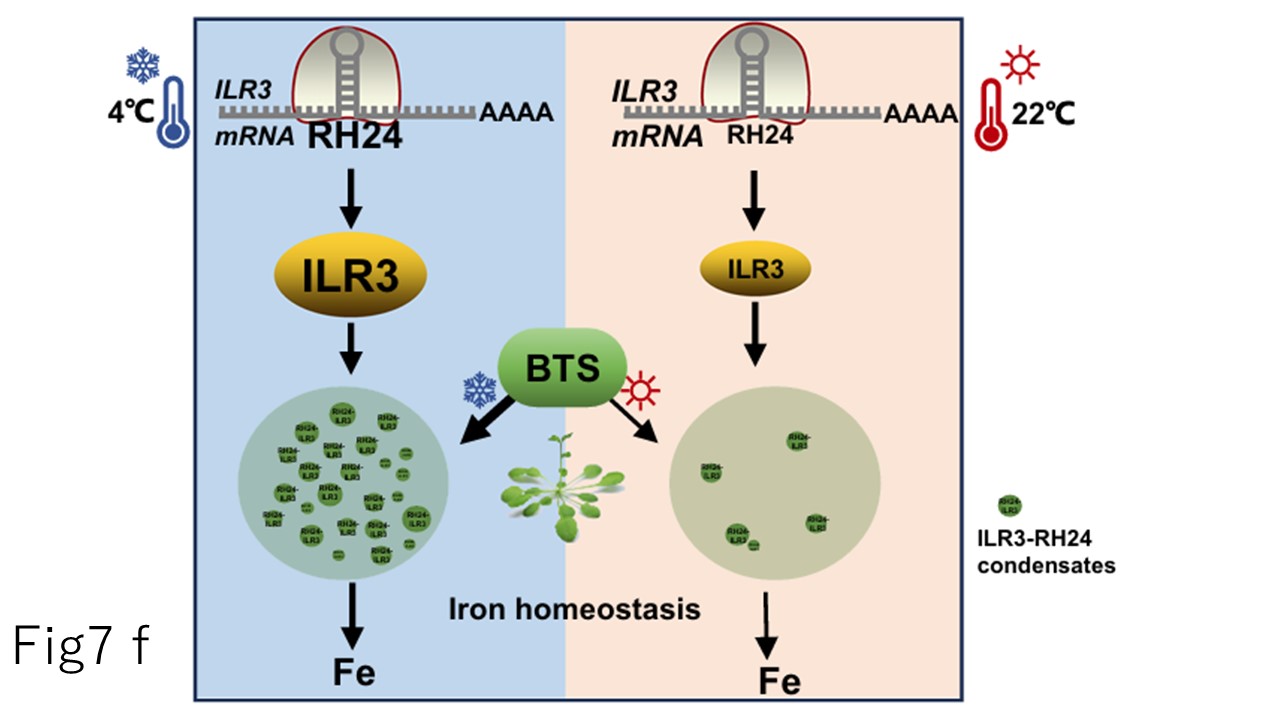
恾俈.f.