僔儘僀僰僫僘僫嬥懏僩儔儞僗僙僾僞乕IRT1儖乕僾偺嬥懏僙儞僒乕摿惈
僔儘僀僰僫僘僫嬥懏僩儔儞僗僙僾僞乕IRT1儖乕僾偺嬥懏僙儞僒乕摿惈
Metal-sensing properties of the disordered loop from the Arabidopsis metal transceptor IRT1
Virginia Cointry, Reyes Ródenas, Nelly Morellet, Steven Fanara, Valérie Cotelle1, Julie Neveu and Grégory Vert
Biochemical Journal (2025) 482 451–466
梫巪
怉暔揝惂屼僩儔儞僗億乕僞乕1乮IRT1乯偼丄揝偑惂尷偝傟偨忦審壓偱崻偵揝傪庢傝崬傓嵶朎枌僞儞僷僋幙偱偁傞丅IRT1偼丄偦偺庡梫側婎幙偱偁傞揝埲奜偵丄搚忞偺揝偑彮側偔IRT1偑崅敪尰偟偰偄傞応崌偵怉暔偵夁忚拁愊偡傞懠偺2壙嬥懏傕桝憲偡傞丅変乆偼丄IRT1偺旕揝嬥懏婎幙偵傛傞東栿屻惂屼偵丄枌娧捠傜偣傫TM4偲TM5偺娫偺嵶朎撪惂屼儖乕僾偑娭梌偟偰偄傞偙偲傪埲慜偵曬崘偟偨丅垷墧偑夁忚偵側傞偲丄IRT1偼CIPK23偵傛傞儕儞巁壔傪庴偗丄懕偄偰IDF1偵傛偭偰儐價僉僠儞壔偝傟丄IRT1偼塼朎暘夝傪庴偗傞丅偙偺垷墧埶懚揑側IRT1偺僟僂儞儗僊儏儗乕僔儑儞偵偼丄IRT1儖乕僾偵垷墧偲捈愙寢崌偡傞4偮偺僸僗僠僕儞乮H乯巆婎偑懚嵼偡傞偙偲偑昁梫偱偁傞丅偟偐偟丄偳偺傛偆偵偟偰慖戰揑側嬥懏寢崌偑払惉偝傟傞偺偐丄傑偨丄偳偺傛偆偵偟偰壓棳偺惂屼偑峴傢傟傞偺偐偼丄傎偲傫偳傢偐偭偰偄側偄丅変乆偼丄IRT1儖乕僾偺嬥懏寢崌摿惈偲峔憿傪柧傜偐偵偟丄旕揝嬥懏僙儞僒乕偲僔僌僫儖揱払偺暘巕婎斦傪傛傝怺偔棟夝偡傞偙偲傪栚巜偟偨丅墌曃岝擇怓惈偲NMR偺慻傒崌傢偣偵傛傝丄垷墧偲儅儞僈儞偑僫僲儌儖椞堟偺恊榓惈偱IRT1儖乕僾偵寢崌偡傞偙偲丄嬥懏寢崌偑儖乕僾偺峔憿壔傪堷偒婲偙偝側偄偙偲傪柧傜偐偵偟偨丅変乆偼丄垷墧偲儅儞僈儞偺寢崌偑4偮偺H巆婎偵傛偭偰攠夘偝傟傞偙偲傪専徹偟丄傾僗僷儔僊儞巁乮D乯巆婎D173偑嬥懏攝埵傪彆偗丄怉暔偵偍偗傞IRT1偺嬥懏姶抦偲嬥懏埶懚惈暘夝偵娭梌偟偰偄傞偙偲傪摨掕偟偨丅埲忋偺寢壥偐傜丄IRT1惂屼儖乕僾偑嵶朎幙撪偺崅擹搙偺擇壙嬥懏傪姶抦偟丄怉暔偵偍偗傞嬥懏庢傝崬傒傪惂屼偟偰偄傞偙偲偑柧傜偐偵側偭偨丅
乮幚尡曽朄偲寢壥偼徣棯乯
峫嶡
曄壔偡傞塰梴嫙媼検偲塰梴廀梫偵懳張偡傞偨傔丄怉暔偼塰梴僩儔儞僗億乕僞乕偺敪尰傪惂屼偡傞愴棯傪奐敪偟偨丅偙傟偼丄僔儘僀僰僫僘僫偺IRT1揝僩儔儞僗億乕僞乕偺応崌偱偁傝丄條乆側嬥懏婎幙偵傛偭偰堎側傞儗儀儖偱惂屼偝傟偰偄傞丅偡側傢偪丄掅Fe2+偵傛傞揮幨惂屼偲丄Zn2+偍傛傃Mn2+夁忚偵傛傞東栿屻惂屼偱偁傞丅埲慜丄変乆偼丄旕揝嬥懏夁忚偵墳摎偟偨嵶朎枌偱偺IRT1偺東栿屻惂屼偵偼丄CIPK23僉僫乕僛偵傛傞儕儞巁壔偲丄偦傟偵懕偔IDF1 E3儕僈乕僛偵傛傞K63億儕儐價僉僠儞嵔偺晅壛偑娭梌偟偰偄傞偙偲傪帵偟偨偑丄偙偺婡峔偵偼IRT1惂屼儖乕僾偺僸僗僠僕儞偵晉傫偩儌僠乕僼偑昁梫偱偁傞丅IRT1偑嬥懏傪姶抦偡傞暘巕婡峔傪偝傜偵怺偔棟夝偡傞偨傔偵丄NMR暘岝朄丄CD暘岝朄丄MST夝愅丄僞儞僷僋幙梊應傾儖僑儕僘儉傪慻傒崌傢偣偰丄IRT1挷愡儖乕僾偺峔憿婎斦傪挷傋偨丅偦偺寢壥丄IRT1儖乕僾偼杮幙揑偵柍拋彉側椞堟乮IDR乯偱偁傝丄條乆側僐儞僼僅儊乕僔儑儞傪偲傞偙偲偑偱偒傞偙偲偑傢偐偭偨丅偙偺摿惈偼丄僸僩偺ZIP4垷墧僩儔儞僗億乕僞乕偵傕尒傜傟傞傛偆偵丄ZIP娫偱曐懚偝傟偰偄傞傛偆偱偁傞丅偟偐偟側偑傜丄僔儘僀僰僫僘僫偺MTP1塼朎垷墧僩儔儞僗億乕僞乕偺傛偆側懠偺僩儔儞僗億乕僞乕僼傽儈儕乕傕丄嬥懏寢崌儌僠乕僼傪帩偮柍拋彉側嵶朎撪儖乕僾傪帩偭偰偄傞偺偱丄偙傟偼ZIP偺摿堎揑側摿挜偱偼側偄丅変乆偺娤嶡偵傛傞偲丄IRT1儖乕僾偼Zn2+偺懚嵼壓偱彫偝側峔憿曄壔傪婲偙偡偑丄偦偺戝晹暘偼柍拋彉側傑傑偱偁傞丅偙偺娤嶡寢壥偼丄hZIP4偱埲慜偵曬崘偝傟偨傛偆偵丄IDR偲彫偝側儕僈儞僪偲偺娫偵乽僼傽僕乕乿暋崌懱偑宍惉偝傟傞偙偲傪帵偟偰偄傞丅杮幙揑偵柍拋彉側僞儞僷僋幙乮IDP乯傗IDR偺憡屳嶌梡偼丄寢崌帪偵姰慡偵僼僅乕儖僨傿儞僌偡傞傕偺偐傜丄偙偺傛偆側乽僼傽僕乕乿暋崌懱傪宍惉偡傞傕偺傑偱丄偝傑偞傑側儊僇僯僘儉傪帵偡偙偲偑偁傞丅乽僼傽僕乕暋崌懱乿偲偼丄娭梌偡傞IDP傗IDR偑寢崌帪偵僐儞僼僅儊乕僔儑儞偺廮擃惈傪曐偮傛偆側憡屳嶌梡偺堦庬傪巜偡丅廬棃偺埨掕側僞儞僷僋幙暋崌懱偲偼堎側傝丄偙傟傜偺憡屳嶌梡偼摦揑側嫇摦傪摿挜偲偟丄暋崌懱偼扨堦偺丄屌掕偝傟偨僐儞僼僅儊乕僔儑儞傪偲傜側偄丅僞儞僷僋幙偵偍偗傞IDR偺廮擃惈偲僐儞僼僅儊乕僔儑儞壜慪惈偑岦忋偡傞偙偲偱丄僷乕僩僫乕娫偺憡屳嶌梡傗壔妛斀墳傪懀恑偡傞東栿屻僀儀儞僩偺偨傔偺僾儔僢僩僼僅乕儉偑採嫙偝傟傞丅IRT1偺応崌丄偙偺傛偆側擻椡偼丄旕揝嬥懏夁忚帪偺IRT1儖乕僾儕儞巁壔偺偨傔偵丄CIPK23僉僫乕僛偺傛偆側壓棳場巕偺儕僋儖乕僩傪懀恑偡傞壜擻惈偑偁傞丅偝傜偵丄IRT1挷愡儖乕僾偺僸僗僠僕儞巆婎偑東栿屻挷愡偵偍偄偰廳梫偱偁傞偙偲偼埲慜偵傕徹柧偝傟偰偍傝丄4偮偺僸僗僠僕儞巆婎傪曄堎偝偣偨僞儞僷僋幙傪帩偮怉暔偼丄旕揝嬥懏夁忚帪偵IRT1偺儕儞巁壔偲暘夝傪帵偝側偄丅嫽枴怺偄偙偲偵丄挷愡儖乕僾偺C枛抂偵埵抲偡傞巆婎偺壔妛僔僼僩偑丄Zn2+懚嵼壓偲旕懚嵼壓偲偱堎側傞偙偲偑娤嶡偝傟偨丅偟偐偟側偑傜丄僸僗僠僕儞偺嫟柭偼Zn2+偺揧壛偵傛偭偰徚幐偡傞偨傔丄偙偺摿掕偺怢挘偑偙偺傛偆側忦審壓偱摿堎揑側僐儞僼僅儊乕僔儑儞傪偲傞偐偳偆偐傪寛掕偡傞偙偲偼偱偒側偐偭偨丅
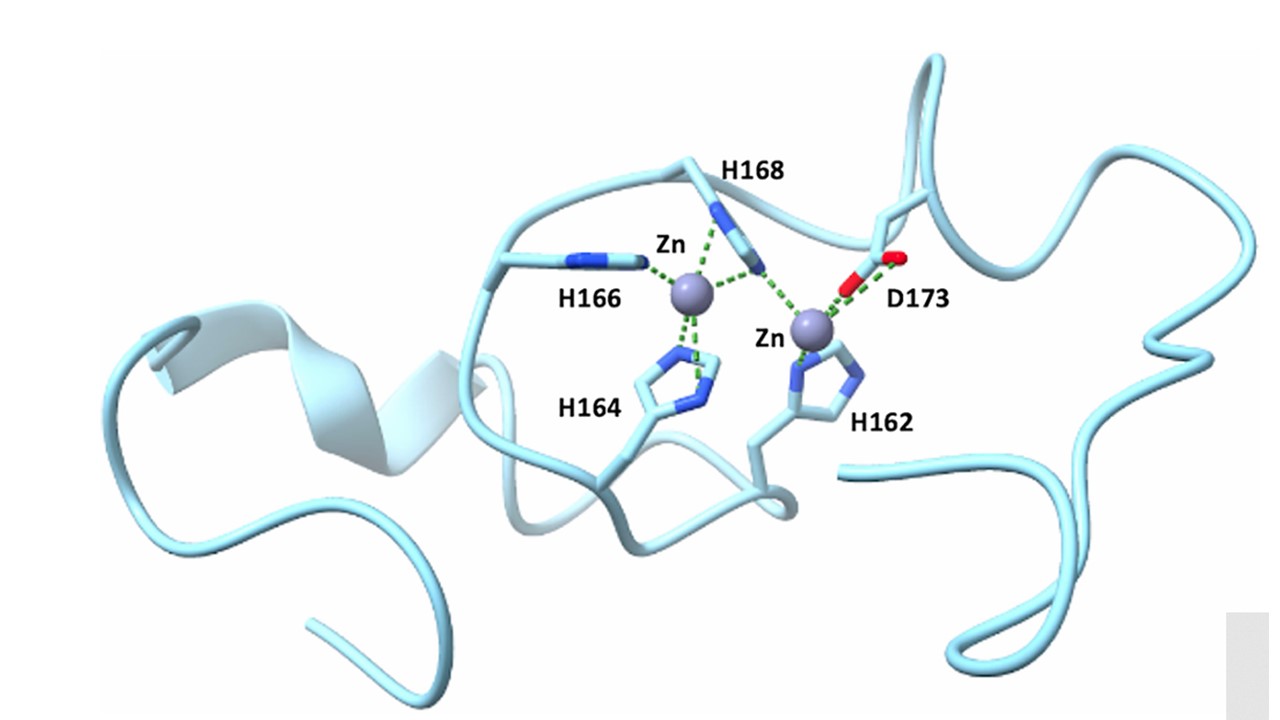
IRT1傊偺Zn2+偲Mn2+偺寢崌偵偮偄偰婰榐偝傟偨僫僲儌儖椞堟偺夝棧掕悢偼丄夁忚忦審壓偱惗堢偟偨怉暔偵偍偄偰幚尡揑偵寛掕偝傟偨僫僲儌儖擹搙偺嬥懏偲堦抳偟偰偄傞丅廳梫側偙偲偼丄僸僗僠僕儞偺怢挘偑丄Zn2+偲Mn2+偺捈愙攝埵偵偲偭偰愨懳揑偵廳梫偱偁傞偙偲傪柧傜偐偵偟偨偙偲偱偁傞丅嬃偔傋偒偙偲偵丄2偮偺僸僗僠僕儞巆婎偑曄堎偟偰傕IRT1儖乕僾偼Zn2+偲寢崌偱偒傞偙偲偑傢偐偭偨丅偍偦傜偔懠偺2偮偺僸僗僠僕儞偲傾僗僷儔僊儞巁173偑攝埵傪扴偭偰偄傞偨傔偱偁傠偆丅偙偺偙偲偼丄僸僗僠僕儞偺擇廳曄堎懱偱摼傜傟偨NMR僨乕僞偐傜傕巟帩偝傟丄巆傝偺2偮偺僸僗僠僕儞巆婎偲傾僗僷儔僊儞巁173偺僾儘僩儞嫟柭偼丄Zn2+懚嵼壓偱偼傗偼傝徚幐偟偨丅Zn2+偺攝埵悢偑3偱偁傞偙偲偼堦斒揑偱偼側偄偑丄Zn2+偼戝偒側僄僱儖僊乕儁僫儖僥傿乕傪庴偗傞偙偲側偔丄條乆側榗傫偩攝埵峔憿傪偲傞偙偲偑婰嵹偝傟偰偄傞丅偦偺忋丄傾僗僷儔僊儞巁巆婎144偵懳偟偰Alphafold僒乕僶乕3偑梊應偟偨傛偆偵丄廃埻偺巆婎偑Zn2+偺攝埵偵婑梌偡傞偙偲偱丄IDR偲偟偰偺IRT1儖乕僾偺廮擃惈偑丄嶰廳曄堎懱H162A/H164A/D173N偺Zn2+寢崌擻偺巆懚傪愢柧偱偒傞偺偱偼側偄偐偲悇應偟偰偄傞乮僆儞儔僀儞曗懌恾10乯丅
NMR偲MST偵傛傞娤嶡偐傜丄IRT1偺挷愡儖乕僾偺5偮偺巆婎偑Zn2+傪攝埵偟偰偄傞壜擻惈偑偁傞丅偡偱偵丄2偮偺Zn2+僀僆儞偑5偮偺僀僈儞僪偵傛偭偰攝埵偝傟丄儕僈儞僪偺1偮偑嫶搉偟偲偟偰摥偔偲曬崘偝傟偰偄傞偺偱丄IRT1儖乕僾偵傕2偮偺Zn2+寢崌晹埵偑懚嵼偡傞壜擻惈偑偁傞丅偙偺壖愢偼丄2偮偺Zn2+僀僆儞偑懚嵼偡傞IRT1儖乕僾偵懳偟偰Alphafold僒乕僶乕3傪梡偄偰峴偭偨梊應偱傕巟帩偝傟偰偍傝丄偦偙偱偼僸僗僠僕儞168偑椉曽偺僀僆儞傪攝埵偡傞嫶偲偟偰帵偝傟偰偄傞乮恾6乯丅巆擮側偑傜丄IRT1儖乕僾偵2偮偺Zn2+寢崌晹埵偑懚嵼偡傞壜擻惈偼丄崱夞偺僨乕僞偱偼嬫暿偱偒側偄丅hZIP4偺僸僗僠僕儞偑朙晉側嵶朎幙儖乕僾偱壖掕偝傟偨傛偆偵丄偍偦傜偔扨堦偺Zn2+寢崌忬懺偼懚嵼偣偢丄傓偟傠Zn2+寢崌忬懺偼丄僸僗僠僕儞巆婎偲傾僗僷儔僊儞巁173偺暋悢偺慻傒崌傢偣偵傛偭偰Zn2+偑攝埵偝傟偨僐儞僼僅儊乕僔儑儞傾儞僒儞僽儖偱偁傞壜擻惈偑崅偔丄嵶朎幙Zn2+擹搙偵墳偠偰IRT1撪偱夁搉揑側Zn2+寢崌條幃傪壜擻偵偡傞偲庡挘偡傞丅
変乆偼傑偨丄Zn2+寢崌偵娭梌偡傞摨偠巆婎偑丄偍偦傜偔椬愙偡傞巆婎偲偲傕偵Mn2+偺攝埵偵傕娭梌偟偰偄傞偙偲傪柧傜偐偵偟偨丅偟偐偟側偑傜丄IRT1儖乕僾偼Mn2+傛傝傕Zn2+偵懳偟偰崅偄恊榓惈傪帵偟丄偙傟偼Zn夁忚偵捈柺偟偨怉暔偱娤嶡偝傟傞IRT1偺嫮偄暘夝偲堦抳偡傞丅IRT1儖乕僾偑Zn2+丄Mn2+丄偁傞偄偼偦偺椉曽傪挷惍偡傞偐偳偆偐偼丄嬥懏僀僆儞偺棙梡壜擻惈偲憡懳揑側嵶朎幙嬊強擹搙偵傛偭偰寛傑傞偲巚傢傟傞丅
慡懱偲偟偰丄変乆偺尋媶偼丄ZIP僩儔儞僗億乕僞乕偺嬥懏姶抦摿惈偺夝愅偺偨傔偺榞慻傒傪採嫙偟丄IRT1僞儞僷僋幙偑偦偺惂屼儖乕僾傪捠偟偰偳偺傛偆偵嬥懏傪姶抦偡傞偺偐偵偮偄偰偺棟夝傪怺傔傞傕偺偱偁傞丅偙偺棟夝偼丄怉暔偑偳偺傛偆偵揝偺庢傝崬傒傪嵟揔壔偟丄怉暔慻怐偵偍偗傞斀墳惈偺崅偄旕揝嬥懏偺媧廂傪惂尷偟偰偄傞偐傪攃埇偟丄怉暔偵偍偗傞廳嬥懏拁愊傪挷愡偡傞僶僀僆僥僋僲儘僕乕揑傾僾儘乕僠傪専摙偡傞忋偱嬌傔偰廳梫偱偁傞丅
恾6偺愢柧丗IRT1挷愡儖乕僾偵傛傞垷墧僀僆儞偲儅儞僈儞僀僆儞偺攝埵丅IRT1儖乕僾偑H162, H164, H166, H168, D173巆婎偲攝埵偡傞Zn2+僀僆儞乮僌儗乕偺媴乯偵懳偡傞Alphafold3偺梊應丅椢怓偺攋慄偼3.5Å埲撪偺嬥懏攝埵傪帵偡丅夝愅偼UScf.ChimeraX傾僾儕働乕僔儑儞偱峴偭偨丅IRT1, 揝惂屼僩儔儞僗億乕僞乕1丅
恾6