夁忚側揝偺拁愊偼丄僨儞僾儞崌惉偺慾奞偲DNA懝彎偺桿敪偵傛偭偰僩僂儌儘僐僔泱擕偺敪払偵塭嬁傪媦傏偡
夁忚側揝偺拁愊偼丄僨儞僾儞崌惉偺慾奞偲DNA懝彎偺桿敪偵傛偭偰僩僂儌儘僐僔泱擕偺敪払偵塭嬁傪媦傏偡
Jie Zang乥Xueyan Yao乥Tengfei Zhang乥Boming Yang乥乥Zhen Zang乥Xueyan Yao乥Tengfei Zhang乥Boming Yang
J Cell Physiol. 2024;e31427.
梫巪
崚暔庬巕拞偺揝乮Fe乯挋憼偼丄僸僩偺庡側怘帠惈揝尮偱偁傞丅僩僂儌儘僐僔乮Zea mays乯偱偼丄庬巕拞偺揝偺拁愊偼廂妌検偲晧偺憡娭偑偁傞偙偲偑抦傜傟偰偄傞丅廬偭偰丄崅廂検偱崚棻拞偺Fe擹搙偑崅偄僩僂儌儘僐僔昳庬傪奐敪丒堢庬偡傞偨傔偵偼丄偦偺婎慴偲側傞儊僇僯僘儉傪棟夝偡傞偙偲偑晄壜寚偱偁傞丅偙偙偱偼丄帋尡娗撪偱偺崚棻攟梴偵惉岟偟丄攟抧拞偺夁忚側揝嫙媼偑崚棻偺曵夡傪堷偒婲偙偟丄崚棻偺怓偑敄偔側傞偙偲傪帵偟偨丅幚嵺丄泱擕拞偺揝偺夁忚拁愊偼ADP-僌儖僐乕僗僺儘儂僗儂儕儔乕僛乮AGPase乯偺懚嵼検偲妶惈傪慾奞偟丄ysl2曄堎懱偱偼AGPase偺彫僒僽儐僯僢僩傪僐乕僪偡傞Briittle 2乮Bt2乯偺夁忚敪尰偵傛偭偰崚棻敪払偺寚懝偑娚榓偝傟偨丅妶惈巁慺庬乮ROS乯偲嵶朎巰偺僀儊乕僕儞僌偲掕検揑夝愅偐傜丄揝僗僩儗僗偑泱擕嵶朎偱ROS僶乕僗僩偲怺崗側DNA懝彎傪桿敪偡傞偙偲偑帵偝傟偨丅偝傜偵丄僀乕僗僩儚儞僴僀僽儕僢僪僗僋儕乕僯儞僌偵傛傝丄崚棻撪偺揝儂儊僆僗僞僔僗偵娭楢偡傞岓曗堚揱巕偲丄ZmYSL2傪惂屼偡傞忋棳偺揮幨場巕傪摨掕偡傞偙偲偵惉岟偟偨丅偙傟傜偺尋媶傪憤崌偡傞偲丄揝偺拁愊偵傛偭偰惂屼偝傟傞庬巕宍惉偺暘巕儊僇僯僘儉偵娭偡傞摯嶡偑摼傜傟丄彨棃揑偵偼僩僂儌儘僐僔偺昳庬夵椙偵偍偗傞揝尦慺偺岠棪揑側棙梡偑懀恑偝傟傞偱偁傠偆丅
乮埲壓杮暥偼3.6偺傒栿偡乯
3.6乥崚棻撪偺揝儂儊僆僗僞僔僗偵娭梌偡傞岓曗堚揱巕偺摨掕
崚棻撪偺揝擹搙偑崅偄僶僀僆嫮壔僩僂儌儘僐僔傪奐敪偡傟偽丄揝寚朢偵傛傞昻寣傪娚榓偱偒傞偙偲偼傛偔抦傜傟偰偄傞丅偟偐偟丄崚棻撪偺崅偄揝擹搙偼僩僂儌儘僐僔偺廂検偲晧偺憡娭偑偁偭偨丅偟偨偑偭偰丄崚棻撪偺揝儂儊僆僗僞僔僗偵婑梌偡傞堚揱帒尮傪摨掕偡傞偙偲偼婱廳偱偁傞丅変乆偼丄暥專乮Gu et al., 2015; Jin et al., 2013; Qin et al., 2012; Zhang et al., 2017乯偐傜丄崚棻撪揝擹搙偵娭楢偡傞 42 偺撈棫偟偨検揑宍幙嵗埵乮QTL乯傪廂廤偟偨丅奺QTL偺僼儔儞僉儞僌儅乕僇乕傪僩僂儌儘僐僔僎僲儉偺暔棟抧恾乮B73 RefGen_v4乯偵傾儔僀儊儞僩偟偨丅偦偟偰丄彮側偔偲傕2偮偺嵞尰惈偺偁傞QTL嬊嵼傪娷傓椞堟傪儂僢僩僗億僢僩椞堟偲掕媊偟丄崚棻Fe擹搙偵偮偄偰12偺QTL儂僢僩僗億僢僩傪摨掕偟偨乮恾7a乯丅僩僂儌儘僐僔崚棻撪偺揝儂儊僆僗僞僔僗偵婑梌偡傞枹抦偺堚揱巕傪扵嶕偡傞偨傔丄惓忢偍傛傃揝夁忚忦審壓偱惗堢偝偣偨崚棻傪梡偄偰RNA-seq傪峴偄丄崚棻偺揝擹搙偵娭楢偡傞尰嵼擖庤壜擻側僩儔儞僗僋儕僾僩乕儉僨乕僞傪摑崌偟偰146偺DEG傪僗僋儕乕僯儞僌偟偨乮恾7b乯乮Yan et al.乯 偝傜偵丄偙傟傜偺 DEGs 偑僩僂儌儘僐僔 10 愼怓懱忋偺 QTL 儂僢僩僗億僢僩撪偵暘晍偟偰偄傞偙偲傪娤嶡偟丄揝僗僩儗僗偵墳摎偡傞偙傟傜偺堚揱巕偺婡擻偵偮偄偰怴偨側抦尒傪摼傞偙偲偑偱偒偨乮恾 7a乯丅
偦偺寢壥丄9屄偺DEG偑儂僢僩僗億僢僩椞堟偵埵抲偟偰偄偨乮恾7a丄Supplementing Information S1丗曗懌僨乕僞乯丅偙傟傜偺堚揱巕偼丄崚棻撪偺揝偺巭寣偵婑梌偟偰偄傞壜擻惈偑偁傝丄偦偺婡擻傪偝傜偵怺 偔挷傋傞昁梫偑偁傞丅
ZmYSL2偼崚棻偺揝儂儊僆僗僞僔僗偵偍偄偰廳梫側僩儔儞僗億乕僞乕婡擻偱偁傞丅偟偨偑偭偰丄ZmYSL2偺敪尰傪惂屼偡傞揮幨場巕偺壜擻惈傪挷傋傞偙偲偼壙抣偑偁傞丅変乆偼丄ZmYSL2僾儘儌乕僞乕偺挿偝偺堎側傞4偮偺抐曅乮Pro1:-1〜-518丄Pro2:-498〜-1007丄Pro3丗 -Pro3: -1484 to -1996 bp乯丄pAbAi儀僋僞乕偵僋儘乕僯儞僌偟丄峺曣僎僲儉偵慻傒崬傫偩丅儔僀僽儔儕乕偺拞偐傜丄6偮偺bHLH揮幨場巕傪娷傓崌寁24偺揮幨場巕偑慖戰偝傟偨丅嫽枴怺偄偙偲偵丄偙傟傜偺bHLH揮幨場巕偼偡傋偰Pro2傪昗揑偲偟偰偄偨丅偦偙偱丄Plant CARE僨乕僞儀乕僗(http://bioinformatics. psb.ugent.be/webtools/plantcare/html/)傪梡偄偰偙偺椞堟偺婛抦偺僔僗僄儗儊儞僩傪専嶕偟偨偲偙傠丄3偮偺CAAT-box儌僠乕僼丄4偮偺MYC儌僠乕僼丄STRE儌僠乕僼丄2偮偺sp1儌僠乕僼偺崌寁10屄偺儌僠乕僼偑摨掕偝傟丄偦偺偆偪MYC儌僠乕僼偼怉暔偺bHLH揮幨場巕偵傛偭偰擣幆偝傟偨丅偦偺屻偺峺曣儚儞僴僀僽儕僢僪幚尡偵傛傝丄偙傟傜6偮偺bHLH揮幨場巕偑in vitro偱妋偐偵ZmYSL2僾儘儌乕僞乕偺儀僀僩攝楍傪擣幆偱偒傞偙偲偑妋擣偝傟偨乮恾7c乯丅偝傜偵丄10杮偺僩僂儌儘僐僔愼怓懱偺暔棟抧恾忋偱24偺揮幨場巕偺暘晍傪娤嶡偟偨丅Zm00001d005100偲Zm00001d048229偼丄偦傟偧傟2斣愼怓懱忋偺158.14-177.06Mb偲9斣愼怓懱忋偺149.61-153.12Mb偺QTL儂僢僩僗億僢僩偵埵抲偟偰偄偨丅Zm00001d042463偲Zm00001d04940偼丄偦傟偧傟戞3愼怓懱偲戞9愼怓懱偺168.69-172.88Mb偲8.83-10.97Mb偺QTL儂僢僩僗億僢僩晅嬤偵埵抲偟偰偄偨乮恾7a乯丅憤崌偡傞偲丄偙傟傜偺揮幨場巕偼僩僂儌儘僐僔崚棻撪偺揝巭寣偵娭梌偡傞僇僕儏傾儖側惂屼場巕偱偁傞壜擻惈偑偁傞丅
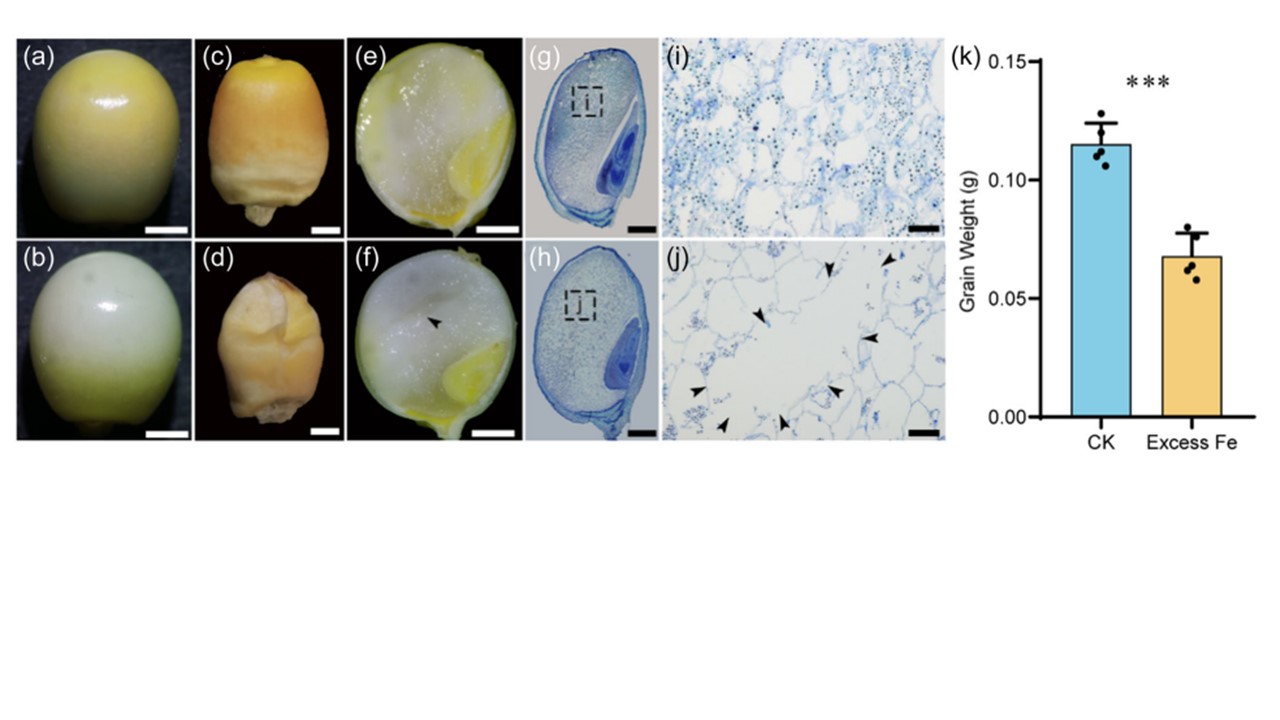
恾 1 惓忢偍傛傃揝夁忚忦審壓偱惗堢偟偨僩僂儌儘僐僔嬤岎宯 B73 偺崚棻偺昞尰宆揑摿挜丅(a, b) 庴暡屻20擔乮DAP乯偺惓忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻丅 (c, d) 惓忢偍傛傃揝夁忚忦審壓偱惗堢偟偨惉弉崚棻偺昞尰宆丅(e, f)庴暡屻20擔栚丄捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻偺廲抐柺丅(g, h) 20 DAP偱捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻偺慻怐妛揑暘愅丅(i, j) 偦傟偧傟(g)偲(h)偺奼戝恾丅(k) 扨堦崚棻偺棻廳偺暘愅丅栴報偼嬻摯傪帵偡丅僄儔乕僶乕偼SD丄Student偺t専掕丟***p < 0.001丅(a-h) scale bar, 1 mm. (i, j)僗働乕儖僶乕丄200兪m丅
恾2
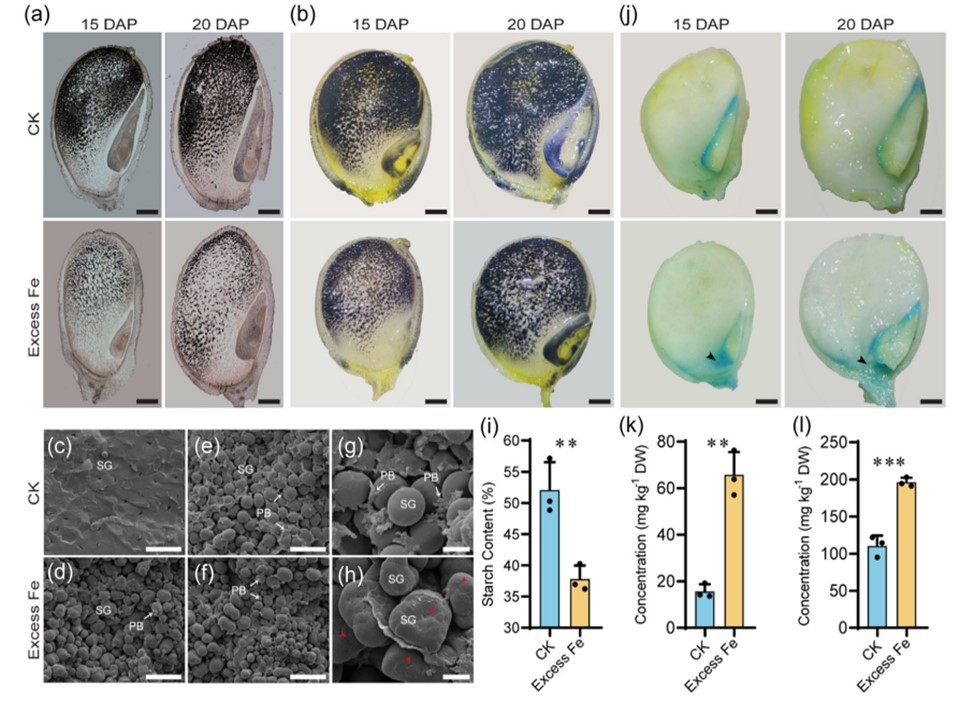
揝夁忚拁愊偼嬤岎宯B73偺崚棻僨儞僾儞娷検偵塭嬁傪梌偊偨丅(a) 捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻偺僷儔僼傿儞愗曅偺儓僂慺愼怓丅(b) 捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨怴慛側崚棻偺儓僂慺愼怓丅(c, d) 惓忢偍傛傃揝夁忚忦審壓偱惗堢偟偨惉弉崚棻偺廃曈泱擕偺憱嵏宆揹巕尠旝嬀(SEM)丅(e, f) 捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨惉弉崚棻偺拞怱晹偺揵暡幙泱擕偺SEM丅(g, h) (e, f)偺旍戝偟偨僨儞僾儞梓棻偺宍懺丅栴報偼奐岥晹傪帵偡丅(i) 惓忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻偺僨儞僾儞娷検丅(j) 僾儖僔傾儞僽儖乕愼怓偵傛傞丄捠忢偍傛傃揝夁忚忦審壓偱惗堢偟偨崚棻偺揝暘晍丅栴報偼揝偺拁愊傪帵偡丅(k) 撪泱擕拞偺揝娷桳検偺應掕丅(l) 泱拞偺揝娷桳検偺應掕丅(a,b偍傛傃j乯僗働乕儖僶乕丄1 mm丅(c-f) scale bar, 20 兪m. (g,h)僗働乕儖僶乕丄4兪m丅僄儔乕僶乕偼SD丄Student's t test; **p < 0.01, **p < 0.001丅PB偼僞儞僷僋幙杮懱丄SG偼僨儞僾儞梓棻丅
恾1
恾2