揮幨場巕Dof3.6/OBP3偼僔儘僀僰僫僘僫偺揝儂儊僆僗僞僔僗傪惂屼偡傞
偙偺尋媶偱偼OBP3偲偄偆Dof偺儂儌儘僌偱偁傞揮幨場巕偑丄嵟廔揑偵揝儂儊僆僗僞僔僗傪惂屼偟偰偄傞偙偲傪敪尒偟偰偄傞丅Dof偼尰嵼搶戝偺惗暔惗嶻岺妛僙儞僞乕偺桍郪廋堦嫵庼偑1990擭戙枛婜偵敪尒偟偨堚揱巕偱偁傞丅
揮幨場巕Dof3.6/OBP3偼僔儘僀僰僫僘僫偺揝儂儊僆僗僞僔僗傪惂屼偡傞
The transcription factor Dof3.6/OBP3 regulates iron homeostasis in Arabidopsis
彊攟攟乮Peipei Xu乯丄梜埶椦乮Yilin Yang乯丄驸拞揤乮Zhongtian Zhao乯丄嬥攷乮Jinbo HH乯丄幱弐墠乮Junyan Xie乯丄墹椡壺乮Lihua Wang乯丄揂宒梇乮Huiqiong Zheng乯丄滹執柧乮Weiming Cai
The EMBO Journal丂13 November 2024
梫巪
揝偼怉暔偵偲偭偰昁恵尦慺偱偁傞丅怉暔偵傛傞揝偺庢傝崬傒偼崅搙偵惂屼偝傟偰偄傞偑丄偦偺崻杮揑側儊僇僯僘儉偼傛偔傢偐偭偰偄側偄丅揝寚朢墳摎惈bHLH100堚揱巕僾儘儌乕僞乕偺愗抐抐曅傪梡偄偰丄僔儘僀僰僫僘僫揮幨場巕峺曣儚儞僴僀僽儕僢僪乮Y1H乯儔僀僽儔儕乕傪僗僋儕乕僯儞僌偟丄DOF僼傽儈儕乕僞儞僷僋幙偱偁傞OBP3偑揝寚朢僔僌僫儖揱払宱楬偺廳梫側峔惉梫慺偱偁傞偙偲傪摨掕偟偨丅OBP3偼丄C枛抂偵妶惈壔僪儊僀儞傪帩偮揮幨梷惂場巕偱偁傞丅偦偺敪尰偼揝寚朢偵傛偭偰桿摫偝傟傞丅OBP3傪夁忚敪尰偝偣偨僩儔儞僗僕僃僯僢僋宯摑偼揝夁忚偲梩偺憗婜夡巰傪帵偟偨偑丄obp3曄堎懱偼揝寚朢偵懳偡傞懴惈偑掅偐偭偨丅OBP3偼丄bHLH堚揱巕僾儘儌乕僞乕偺Ib僒僽僌儖乕僾傪捈愙昗揑偲偡傞偙偲偑敪尒偝傟偨丅OBP3偼bHLH揮幨場巕偱偁傞ILR3乮IAA-LEUCINE RESISTANT3乯偲憡屳嶌梡偟丄偦偺憡屳嶌梡偵傛傝OBP3偺DNA寢崌擻偲揮幨懀恑妶惈偑憹嫮偝傟丄揝寚朢墳摎堚揱巕傪億僕僥傿僽偵惂屼偡傞偙偲偑傢偐偭偨丅偝傜偵丄E3儕僈乕僛BRUTUS偼丄26S僾儘僥傾僜乕儉傪夘偟偨OBP3僞儞僷僋幙偺暘夝傪懀恑偟丄怉暔偵偍偗傞夁忚側揝偺庢傝崬傒傪杊偄偱偄傞丅寢榑偲偟偰丄変乆偺尋媶偼丄怉暔偺揝儂儊僆僗僞僔僗偺惂屼偵偍偗傞OBP3偺廳梫側栶妱傪嫮挷偟偰偄傞丅
峫嶡
Ib僒僽僌儖乕僾偺bHLH堚揱巕偑僔儘僀僰僫僘僫偺揝寚朢墳摎偵廳梫側栶妱傪壥偨偡偙偲偼偡偱偵曬崘偝傟偰偄偨乮Sivitz et al, 2012乯丅変乆偼丄bHLH100堚揱巕僾儘儌乕僞乕傪儀僀僩偲偟偰峺曣儚儞僴僀僽儕僢僪僗僋儕乕僯儞僌傪峴偄丄堚揱巕忋棳偺揮幨場巕傪摨掕偟偨丅僗僋儕乕僯儞僌偺寢壥丄懡偔偺梲惈僐儘僯乕偑摼傜傟丄偦偺偆偪偺7偮偼婛抦偺IVc僒僽僌儖乕僾偺bHLH堚揱巕偵懏偡傞bHLH100偺僾儘儌乕僞乕偵寢崌偟偨丅懠偺2偮偺梲惈僋儘乕儞偼丄DOF僼傽儈儕乕揮幨場巕OBP3偱偁傞偙偲偑傢偐偭偨丅偝傜側傞幚尡偵傛傝丄OBP3偲ILR3偺椉曽偑bHLH100僾儘儌乕僞乕傪妶惈壔偡傞偙偲偑妋擣偝傟偨乮恾1乯丅BP3偼偝傜偵丄in vitro偱偺DNA偍傛傃僞儞僷僋幙寢崌摿惈傪昡壙偡傞偙偲偵傛偭偰摿挜偯偗傜傟偨丅偙偺尋媶偱偼丄OBP傪戝挵嬠偱敪尰偝偣偨丅恾1D偵帵偡傛偆偵丄OBP偼bHLH100僾儘儌乕僞乕偺A/TAAAG僄儗儊儞僩偵寢崌偟偨丅奺AAAG儌僠乕僼偵2bp偺曄堎傪娷傓曄堎宆AAAG僄儗儊儞僩偵偼偳偺OBP傕寢崌偣偢丄偙偺僄儗儊儞僩偵懳偡傞摿堎惈偑帵偝傟偨乮恾6A,B乯丅夁嫀偺暥專偵傛傞偲丄OBP偼僔儘僀僰僫僘僫GST6僾儘儌乕僞乕偺CTTT[T/A]僄儗儊儞僩偵傕摨條偵寢崌偱偒傞乮Kang et al, 2003; Kang and Singh, 2000乯丅
怉暔摿堎揑僕儞僋僼傿儞僈乕僞儞僷僋幙僼傽儈儕乕偵偼丄摿掕偺bZIP僞儞僷僋幙丄崅堏摦搙僌儖乕僾乮HMG乯僞儞僷僋幙偍傛傃懠偺DOF僞儞僷僋幙偲憡屳嶌梡偡傞DOF僞儞僷僋幙偑娷傑傟傞乮Yanagisawa, 2002乯丅OBP僞儞僷僋幙娫偺憡摨惈偼52傾儈僲巁偺DOF寢崌僪儊僀儞偵尷傜傟偰偄傞丅偙傟偑桞堦偺嫟捠攝楍偱偁傞偙偲偐傜丄偙偺僪儊僀儞偼DNA寢崌傗僞儞僷僋幙娫憡屳嶌梡側偳丄暋悢偺婡擻傪帩偮壜擻惈偑偁傞丅OBP僞儞僷僋幙偺DNA寢崌摿惈偼in vitro偱椶帡偟偰偍傝丄bZIP揮幨場巕OBF4偲憡屳嶌梡偡傞偙偲偑偱偒傞丅偙偺寢壥偼丄僩僂儌儘僐僔DOF僞儞僷僋幙偺DNA寢崌偲HMG僞儞僷僋幙偲偺憡屳嶌梡偵DOF僪儊僀儞偑昁恵偱偁傞偲偄偆Yanagisawa (1997)偺抦尒偲堦抳偟偰偄傞(Yanagisawa, 2002)丅僕儞僋僼傿儞僈乕僪儊僀儞偼懠偺僞儞僷僋幙偱傕懡婡擻偱偁傞偙偲偑帵偝傟偰偄傞丅桍郪偵傛傞偲丄僩僂儌儘僐僔DOF1僞儞僷僋幙偺僩儔儞僗妶惈壔妶惈偼丄僉僒儞僩僎儞壔僾儘僩僾儔僗僩偱偼慾奞偝傟偨偑丄椢怓僾儘僩僾儔僗僩偱偼慾奞偝傟側偐偭偨(Yanagisawa, 2000)丅僩僂儌儘僐僔PBF偼bZIP僞儞僷僋幙Opaque2偲憡屳嶌梡偡傞乮Zhan et al, 2018乯丅僞儞僷僋幙娫偺憡屳嶌梡偼摿堎惈偵傕婑梌偟丄慻怐摿堎揑側敪尰僷僞乕儞偵傛偭偰偝傜偵憹嫮偝傟丄慻怐偛偲偵堎側傞揮幨僾儘僼傽僀儖傪傕偨傜偡丅
傑偨丄僩僂儌儘僐僔DOF1僞儞僷僋幙偑僩僂儌儘僐僔C4儂僗儂僄僲儔乕僛僾儘儌乕僞乕偺暋悢偺DOF寢崌晹埵傪夘偟偰僩儔儞僗妶惈壔場巕偲偟偰摥偔偺偵懳偟丄僩僂儌儘僐僔DOF2僞儞僷僋幙偼DOF1僩儔儞僗妶惈壔偺梷惂場巕偲偟偰婡擻偡傞偙偲偑桍郪偵傛偭偰帵偝傟偰偄傞乮Yanagisawa, 2000, 2002乯丅OBP3僞儞僷僋幙偼丄暋悢偺DOF寢崌儌僠乕僼傪娷傓壓棳堚揱巕僾儘儌乕僞乕傪帩偮儗億乕僞乕堚揱巕傪桳堄偵僩儔儞僗妶惈壔偡傞偙偲偼娤嶡偝傟側偐偭偨丅偙偺寢壥偵偮偄偰偼丄OBP3偑揮幨妶惈壔場巕偲偟偰偱偼側偔丄揮幨慾奞場巕偲偟偰摥偄偨壜擻惈側偳丄偄偔偮偐偺愢柧偑偱偒傞丅偟偐偟側偑傜丄GAL4 DNA寢崌僪儊僀儞偲梈崌偟偨OBP偺N枛抂傪寚幐偝偣偨傕偺偼揮幨妶惈傪帵偟偨偙偲偐傜丄OBP偼儔儞妶惈壔僪儊僀儞傪娷傫偱偄傞偙偲偑帵偝傟偨丅偙傟傜偺椞堟偵偼丄僩僂儌儘僐僔DOF1乮Cavalar et al, 2007乯偱娤嶡偝傟偨傛偆側丄婛抦偺妶惈壔僪儊僀儞偲偺憡摨惈偼妋擣偝傟側偐偭偨丅OBP3傪夁忚敪尰偝偣偨僩儔儞僗僕僃僯僢僋怉暔偵傛傝丄偙偺僞儞僷僋幙偑怉暔偺敪惗偵偍偄偰廳梫側栶妱傪壥偨偟偰偄傞偙偲偑埲慜偵帵偝傟偰偄傞乮Kang et al, 2003乯偑丄偙偺堚揱巕偺恀偺婡擻偼埶慠偲偟偰晄柧偱偁傞丅崱夞偺尋媶偱偼丄OBP3偑揝儂儊僆僗僞僔僗偺廳梫側惂屼場巕偱偁傞偙偲傪柧傜偐偵偟偨乮恾9乯丅
偝傜偵丄OBP3夁忚敪尰偵傛傞夁忚揝偺拁愊偲揝儂儊僆僗僞僔僗偺僶儔儞僗偑丄35S::OBP3 lant偺梩偺僋儘儘僔僗偲徕惈昞尰宆偺庡側尨場偱偁傞偙偲傪柧傜偐偵偟偨乮恾2乯丅偝傜偵丄偙偺曄堎昪偼丄廫暘側揝暘傪娷傓1/2MS攟抧偱偼椙岲偵惗堢偟偨偑丄揝寚朢攟抧傗揝偺惗暔妛揑棙梡擻偺掅偄傾儖僇儕惈搚忞偱偼堎忢側惗堢傪帵偟偨偙偲偐傜丄OBP3偑揝偺儂儊僆僗僞僔僗惂屼偵娭梌偟偰偄傞偙偲偑帵偝傟偨丅OBP3偼DOF擣幆攝楍[A/T] AAG偵摿堎揑偵寢崌偟丄揮幨傪妶惈壔偡傞乮Kang et al, 2003乯丅変乆偼丄OBP3偑庡梫側揝儂儊僆僗僞僔僗1b僒僽僌儖乕僾偺bHLH堚揱巕偲怉暔偺惉挿傪億僕僥傿僽偵惂屼偟偰偄傞偙偲傪敪尒偟偨乮恾4乯丅
偙偺敪尒偼丄揝寚朢娐嫬壓偱揝摨壔偺惓妋側憌暿惂屼傪僔僌僫儖偡傞揝寚朢僇僗働乕僪偑丄偙傟傑偱偺娤嶡寢壥偲堦抳偟偰偄傞丅obp3偍傛傃ilr3曄堎懱偺崻偺昞尰宆偼丄揝偑廫暘側忦審壓偱偼栰惗宆偲椶帡偟偰偄傞偑丄掅揝忦審壓偱偼寚懝偟偰偄傞偲偄偆娤嶡寢壥偼丄偙傟傜2偮偺挷愡堚揱巕偑揝寚朢僔僌僫儖揱払宱楬偵偍偄偰摿堎揑側栶妱傪壥偨偟偰偄傞偙偲傪帵嵈偟偰偄傞丅OBP3寢崌嫮搙偼丄bHLH100傪娷傓ILR3偑懚嵼偡傞応崌丄in vivo偱憹壛偡傞乮恾6D乯丅偝傜偵in vitro偱偼丄ILR3bHLH偑寢崌偟偨寢壥丄OBP3DOF偼DOF寢崌晹埵偺椉曽偑曄堎偟偨DNA抐曅偵寢崌偡傞丅偙偺偙偲偼丄DNA偺傾儘僗僥儕乕偑丄ILR3bHLH-DNA暋崌懱偵懳偡傞OBP3DOF偺恊榓惈傪崅傔偰偄傞壜擻惈傪帵嵈偟偰偄傞丅TF偼丄摿掕偺墫婎攝楍偲偺捈愙揑側憡屳嶌梡偲丄DNA偺孅嬋傗姫偒栠偟偲偄偭偨嬊強揑側DNA偺摿挜傪擣幆偡傞偙偲偺椉曽偵傛偭偰丄DNA寢崌晹埵傪擣幆偡傞丅傢傟傢傟偼丄ILR3偑DNA寢崌傪憹嫮偟丄DNA偺嬊強揑側宍忬傪曄壔偝偣傞偙偲偱丄OBP3偺DNA傊偺寢崌偑憹嫮偝傟傞偲峫偊偰偄傞丅OBP3偑惗懱撪偱ILR3偺寢崌偵塭嬁傪媦傏偡偐偳偆偐偵偮偄偰偼丄幚尡傪峴偆昁梫偑偁傞丅bHLH僼傽儈儕乕TF偼巐検懱傪宍惉偡傞偙偲偑帵偝傟偰偄傞丅偙傟傜偺巐検懱偼丄PIF4巐検懱乮Gao et al, 2022b乯偱帵偝傟偰偄傞傛偆偵丄DNA儖乕僾宍惉傪懀恑偟丄嬤朤偺OBP3寢崌傪憹壛偝偣傞壜擻惈偑偁傞丅ILR3偲OBP3僞儞僷僋幙偺憡屳嶌梡偼丄婡擻揑側僿僥儘擇検懱偺宍惉偵偮側偑傞壜擻惈偑偁傝丄偙偺僿僥儘擇検懱偼掅揝忦審壓偱拁愊偟丄揝寚朢壓偱偼庢傝崬傒傪懀恑偡傞丅
BTS傪夘偟偨儐價僉僠儞壔偑OBP3偺暘夝偵偮側偑傞偐偳偆偐傪昡壙偡傞偨傔丄僔儘僀僰僫僘僫偺僾儘僩僾儔僗僩偵BTS-GFP偲Flag-OBP3傪嫟摫擖偟偨丅Flag-OBP3僞儞僷僋幙儗儀儖偼BTS-GFP偲偲傕偵桳堄偵尭彮偟偨偑丄敪尰儗儀儖偼尭彮偟側偐偭偨丅26S僾儘僥傾僜乕儉慾奞嵻偱偁傞50兪M MG132偱張棟偡傞偲丄FlagOBP3儗儀儖偼夞暅偟偨乮晅榐恾S9乯丅偙偺偙偲偼丄BTS1偑in vivo偵偍偄偰26S僾儘僥傾僜乕儉宱楬傪夘偟偨OBP3偺暘夝傪懀恑偡傞偙偲傪帵偟偰偄傞丅偮傑傝杮尋媶偼丄BTS偑26S僾儘僥傾僜乕儉傪夘偡傞DOF揮幨場巕OBP3偲bHLH揮幨場巕ILR3傪摿堎揑偵昗揑偲偟丄偦傟偵傛偭偰ILR3偲OBP3僞儞僷僋幙偑娭梌偡傞僿僥儘擇検懱暋崌懱偺宍惉傪慾奞偡傞偙偲傪帵偡宱尡揑徹嫆傪採帵偟偨丅偦偺寢壥丄偙偺姳徛偼丄揝寚朢墳摎堚揱巕偑惂屼偡傞壓棳偺揝寚朢墳摎堚揱巕偺惂屼傪棎偡壜擻惈偑偁傞丅偝傜偵丄BTS傕揝寚朢忬懺偱偼拁愊偟丄OBP3/ILR3傪夘偟偨墳摎傪慾奞偡傞壜擻惈偑偁傞丅偙偺傛偆偵揝寚朢墳摎傪旝挷惍偡傞偙偲偱丄夁忚偵揝傪庢傝崬傓偙偲側偔丄揔愗側揝媧廂傪曐徹偟丄寢壥偲偟偰崻慡懱偺惉挿偵塭嬁傪梌偊傞丅慡懱偲偟偰丄崱夞偺寢壥偼丄揝寚朢偑偳偺傛偆偵怉暔堚揱巕偺敪尰傪惂屼偟丄偦傟偑偳偺傛偆偵惓忢側惉挿偵婑梌偡傞偺偐偵偮偄偰偺棟夝傪怺傔傞偙偲偵峷專偡傞傕偺偱偁傞丅
恾9.
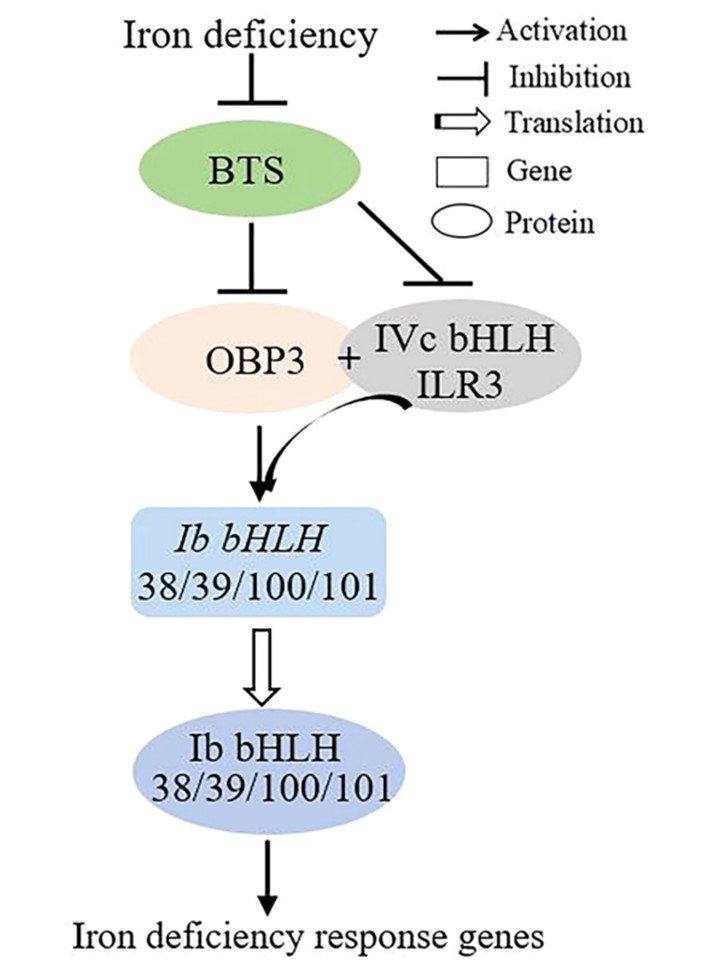
OBP3偑僔儘僀僰僫僘僫偺揝儂儊僆僗僞僔僗傪挷愡偡傞嶌嬈儌僨儖丅
偙偺儌僨儖偼丄僔儘僀僰僫僘僫偺揝儂儊僆僗僞僔僗偵偍偗傞OBP3偲ILR3偺暘巕婡擻傪帵偟偰偄傞丅OBP3偼ILR3偲憡屳嶌梡偟丄Ib僒僽僌儖乕僾bHLH堚揱巕偺僾儘儌乕僞乕偵寢崌偟偰敪尰傪惂屼偡傞丅OBP3偼LR3(IAA-LEUCINE RESISTANT3)偲憡屳嶌梡偟丄偦偺憡屳嶌梡偵傛偭偰OBP3偺DNA寢崌擻偲揮幨懀恑妶惈偑憹嫮偝傟丄偦偺寢壥丄揝寚朢墳摎堚揱巕偺敪尰偑億僕僥傿僽偵惂屼偝傟傞丅揮幨屻儗儀儖偱偼丄E3儕僈乕僛BRUTUS (BTS)偑26S僾儘僥傾僜乕儉婡峔偵傛傞OBP3偲ILR3僞儞僷僋幙偺暘夝傪曗彆偟丄夁忚側揝偺庢傝崬傒傪杊偓丄怉暔偵偍偗傞揝偺峆忢惈傪堐帩偟偰偄傞丅拞怱揑側惂屼僞儞僷僋幙傪懭墌偱丄壓棳偺昗揑堚揱巕傪巐妏偱昞偡丅