鉄栄養に光を当てる:光と鉄欠乏症シグナルにおける転写因子カスケードの交差を探る
鉄栄養に光を当てる:光と鉄欠乏症シグナルにおける転写因子カスケードの交差を探る
Shedding light on iron nutrition: exploring intersections of transcription factor cascades in light and iron deficiency signaling
Ksenia Trofmov, Samriti Mankotia, Mary Ngigi, Dibin Baby, Santosh B. Satbhai and Petra Bauer
Journal of Experimental Botany https://doi.org/10.1093/jxb/erae324
要旨
植物のダイナミックな環境では、光に依存した成長と鉄栄養の相互作用が繰り返し課題となる。植物は、根圏からの鉄の獲得と内部鉄プールの再配置を促進することで、成長と生理を調整し、低鉄レベルに応答する。鉄欠乏応答アッセイと遺伝子共発現ネットワークは、生理学的反応を記録し、遺伝子制御カスケードを解明するのに役立ち、ホルモンと外部シグナル伝達経路の相互作用についての洞察を提供する。しかし、鉄栄養における光の重要性を直接的に探る研究は、まだ限られている。本総説では、鉄欠乏症の制御とその明瞭な光シグナルとの相互関連について、転写因子カスケードと長距離シグナル伝達に焦点を当てて概観する。概日時計と逆行性シグナル伝達は、鉄の取り込みと配分に影響を与える。光によって活性化されるシュートから根への移動性転写因子ELONGATED HYPOCOTYL5(HY5)は、根における鉄ホメオスタシス応答に影響を与える。青色光は、鉄欠乏症を誘発するタンパク質複合体を含む生体分子凝縮体の形成を誘発する。光と鉄シグナル伝達の関連を利用する可能性は、まだ十分に活用されていない。気候変動と土壌のアルカリ性化が進む中、養分利用効率を改善し、光依存性を改変した作物を開発する必要がある。光シグナル伝達と鉄の相互作用を理解し、活用するためには、さらなる研究が必要である。
結論
光は、栄養の利用可能性に加えて、植物がその変化を直接感知し、それに応じて成長を方向転換させる主要な環境的手がかりである。以上の分析から、光と鉄栄養シグナル伝達は複数の点で相互に関連していることがわかった。遺伝子を「赤」、「青」、「バラ」、「鉄分充足」遺伝子に細分化し、それらの制御原理を理解するというコンセプトは、鉄栄養に対する遺伝的・環境的影響と結果を定義するのに役立つ。鉄と光のシグナル伝達における相互関係をよりよく理解するために、今後の研究が強化されることは明らかである。将来的には、以下のようなまだ研究されていない未解決の問題に取り組むことができるだろう。- 低鉄分条件下で蒸散はどのように制御されているのか?鉄の利用可能性は、生物的ストレスに対する鉄栄養を調整するための維管束の分化にどのように影響するか?- 落葉は鉄の獲得にどのように影響するのか?落葉は、植物発生初期に自然に起こることもあれば、植物成長中の光環境の変化の結果として現れることもある。落葉した器官と落葉していない器官では、鉄の吸収源としてどのような能力があるのか?鉄の供給は、落葉した葉と落葉していない葉の鉄獲得能力にどのような影響を及ぼすか?どのシグナルが関与しているのか?- 成長期における光条件、光の方向、遮光、光強度の変化は、植物の鉄利用能力にどのような影響を与えるか?鉄の利用可能性は、生物学的ストレスに鉄栄養を調整するための血管の分化にどのように影響するか?- 落葉は鉄の獲得にどのように影響するのか?落葉は植物発生初期に自然に起こることもあれば、植物成長中の光環境の変化の結果として現れることもある。落葉した器官と落葉していない器官では、鉄の吸収源としてどのような能力があるのか?鉄の供給は、落葉した葉と落葉していない葉の鉄獲得能力にどのような影響を及ぼすか?どのシグナルが関与しているのか?- 成長期における光条件、光の方向、遮光、光強度の変化は、植物の鉄利用能力にどのような影響を与えるか?- 鉄のシグナル伝達によって、内部の鉄供給はどのように制御されるのか?どの感知メカニズムが重要なのか?葉緑体を介した逆行性シグナル伝達とオートファジーの役割は?これらのプロセスは、強光調節やエチレンなどの植物ホルモンの作用とどのように関連しているのか?一酸化窒素やヘムなど、他のシグナル伝達分子はここに関連しうるのか?- 全身的な葉の鉄シグナルは、光とどのように関係しているのか?全身的なシグナルの性質とはどのようなものだろうか?- HY5シグナルは鉄欠乏応答にどのように特異性を与えるのか?HY5シグナルは鉄獲得を制御するbHLHシグナルとどのように相互作用するのか?HY5は根のどこで効果を発揮しているのか?- 鉄シグナル伝達におけるFIT核小体の機能と、どのような条件がその出現を促進するのか。鉄欠乏症応答遺伝子のalternative splicingの役割は何か?FITは根で光シグナル伝達成分とタンパク質相互作用を形成しているのか?他の鉄欠乏症応答制御因子も、液液相分離後の核下凝縮体に蓄積しているのか?地球の気温が上昇し、農業目的の土壌がますますアルカリ性になる中、我々は新しい作物を試し、設計すべきである。そのためには、新規作物が新しい環境でもライフサイクルを完結できるように、養分利用特性を改善し、光周期依存性を修正する必要があるだろう。これまでのところ、異なる土壌における光周期の変化が鉄栄養に及ぼす影響は、作物の改良においてほとんど考慮されていない。栄養価の高い持続可能な作物を開発する必要があるため、光シグナル伝達と鉄栄養の相互関係を理解し利用するために、より多くの研究を行うべきである。
以下図の説明(図2、図3、図4の実を訳した)
図2. 植物の鉄ホメオスタシスを制御するbHLH転写因子カスケード。植物の鉄ホメオスタシス経路は、bHLH転写因子、E3リガーゼ、およびその他のタンパク質によって厳密に制御されている。(A)BTS(BRUTUS)タンパク質やBTSL(BTS-like)タンパク質などのE3リガーゼは、bHLH IVc転写因子をプロテアソーム分解の標的とすることにより、そのタンパク質レベルを制御することができる。鉄欠乏時には、IMA(IRON MAN)タンパク質がBTS/Lと直接結合し、分解の代替標的として機能することにより、BTS/Lの機能を抑制し、bHLH IVcタンパク質の存在量を増加させる可能性がある。(B)鉄欠乏応答につながる転写カスケード。bHLH IVcとIVb転写因子の相互作用は、別のbHLHサブグループIVbメンバーであるPYE(POPEYE)とbHLHサブグループIb(bHLH38、bHLH39、bHLH100、bHLH101)の転写制御のための下流ターゲティングにつながる。例えば、bHLH IVcとURI(UPSTREAM REGULATOR OF IRT1)/bHLH121は活性化につながり、bHLH IVcとbHLH11は非活性化につながる。活性型PYEタンパク質は、特定の標的遺伝子、FRO3 (FERRIC REDUCTASE OXIDASE3)、NAS4 (NICOTIANAMINE SYNTHASE4)、ZIF1 (ZINC FACILITATOR1)、BHLH IBの発現を抑制し、鉄の再分配を引き起こす。一方、bHLH Ib転写因子は、中心的な制御因子であるFIT (FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR)とヘテロ二量体化することにより、根の鉄獲得を活性化し、根誘導遺伝子FRO2 (FERRIC REDUCTASE OXIDASE2)とIRT1 (IRON-REGULATED TRANSPORTER1)を活性化する。破線は、bHLH IVcおよびIVbによる標的遺伝子の制御も可能であることを示している。カラーコードは、遺伝子制御の明確なパターンを示す:「青色」は、鉄欠乏により誘導され、FITに依存し、主に苗の根で作用する;「赤色」は、鉄欠乏により誘導されるが、FITに依存せず、苗の根と芽で作用する;「バラ色」は、鉄欠乏により誘導されず、FITに依存せず、苗の根と芽で作用する。
図3. 鉄ホメオスタシスの光依存的制御。異なる光条件や暗さは、黒い矢印で示すように、鉄の獲得と配分にプラスまたはマイナスの影響を与える。赤色光と青色光は、暗闇や遠赤色光とは対照的に、緑色と赤色の矢印で示すように、遺伝子をアップレギュレートしたりダウンレギュレートしたりする。鉄欠乏成分のカラーコードは図2の通り。概日時計は鉄の恒常性を制御している。鉄が欠乏すると概日周期が長くなる。概日時計成分であるCCA1(CIRCADIAN CLOCK ASSOCIATED1)とTIC(TIME FOR COFFEE)は、鉄欠乏応答遺伝子や鉄充足マーカーFER1(FERRITIN1)のプロモーターに直接干渉する可能性がある。葉緑体、高照度、エチレンにおける逆行性シグナル伝達が交錯し、鉄ホメオスタシスを制御しているのかもしれない。
図4. 鉄の状態の長距離シグナル伝達、多様な全身的鉄シグナルの存在を示す。(A-C)全身性のシュートから根への鉄シグナルの重要性を明らかにするために採用した3つの方法の例。(A)エンドウ(Pisum sativum)のbrz(ブロンズ)やdgl(退化葉状体)などの葉の鉄過剰蓄積変異体を用いた互い接ぎは、根への鉄の取り込みと葉への鉄の配分を促進する構成的な長距離シュートから根への鉄欠乏シグナルの存在を示した。(B)鉄欠乏植物への葉面鉄施用は、鉄充足シグナルによる根への鉄取り込み応答のダウンレギュレーションを引き起こす。しかしながら、葉面散布は、葉茎における長距離鉄充足シグナルの伝達に関与する成分であるOLIGOPEPTIDE TRANSPORTER3が欠損したエンドウ豆dglおよびシロイヌナズナopt3変異体を救済しない。(C) スプリットルートアッセイにより、鉄に曝露された根の半分では局所的・全身的な鉄欠乏シグナルに応答して鉄の取り込みが増加するが、鉄欠乏状態に維持された根の半分では鉄の取り込みが増加しないことが示された。(D) HY5 (ELONGATED HYPOCOTYL5)は、根における鉄ホメオスタシスの制御因子である。光は、例えば赤色光や青色光に応答して、様々な光受容体を活性化させる。活性化した光受容体はCOP1(CONSTITUTIVE PHOTOMORPHOGENIC1)-SPA(SUPRESSOR OF PHYTOCHROME A)複合体を阻害し、HY5の分解を防ぐ。HY5(ELONGATED HYPOCOTYL5)は、根における鉄のホメオスタシスを制御する。光は、例えば赤色光や青色光に反応して、様々な光受容体を活性化させる。活性化した光受容体はCOP1(CONSTITUTIVE PHOTOMORPHOGENIC1)-SPA(SUPRESSOR OF PHYTOCHROME A)複合体を阻害し、HY5の分解を防ぐ。HY5の小さなプールはSPAの活性下でリン酸化されたままであり、光の下でも不活性である。PPK1はHY5に作用する光制御タンパク質キナーゼである。A.thaliana、トマト(Solanum lycopersicum)、リンゴ(Malus baccata)での研究で示されたように、活性のある非リン酸化HY5は光条件下で蓄積し、鉄に関連する様々な応答を誘導する根の移動性シグナルとして働く。A. thalianaでは、HY5は鉄の獲得と配分のためのいくつかの遺伝子の発現を直接制御することができる。あるいは、HY5はホルモンと銅のホメオスタシスにおける機能の結果として、間接的に鉄のホメオスタシスを制御する可能性もある。トマトでは、HY5はFITのオルソログであるFERの作用を妨害することができる。リンゴでは、HY5はYELLOWSTRIPE-LIKEトランスポーター遺伝子(YSL7)に作用する。
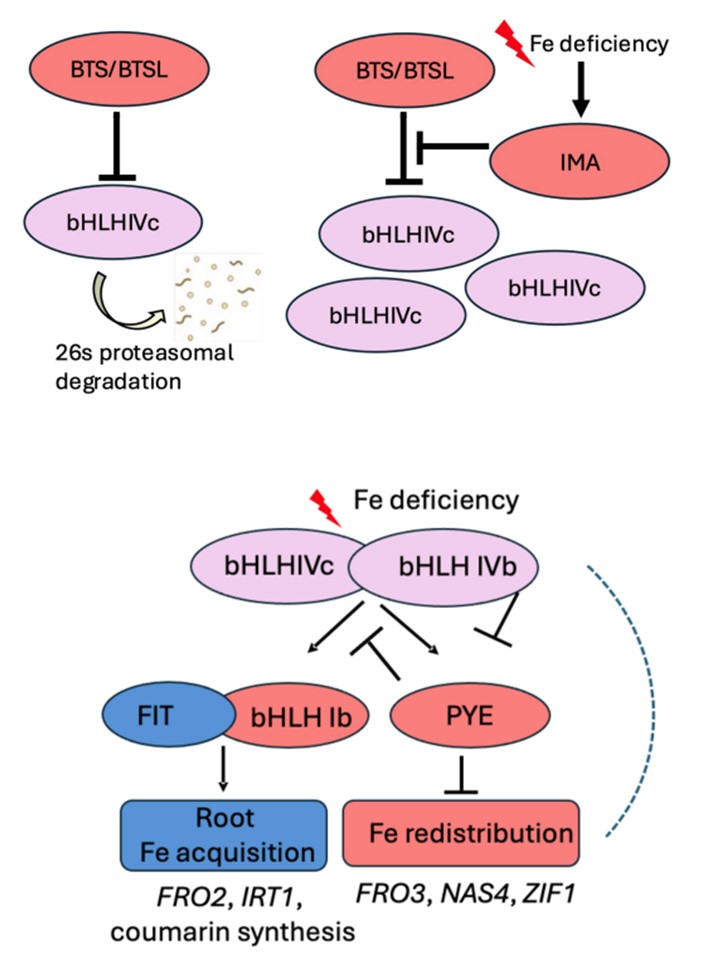
図2
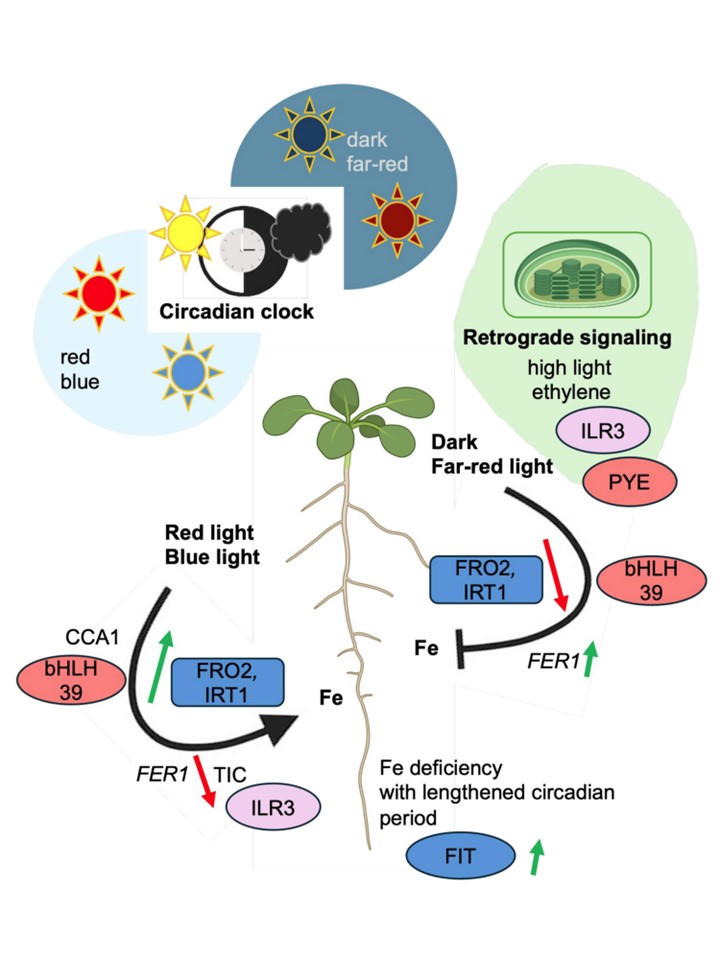
図3
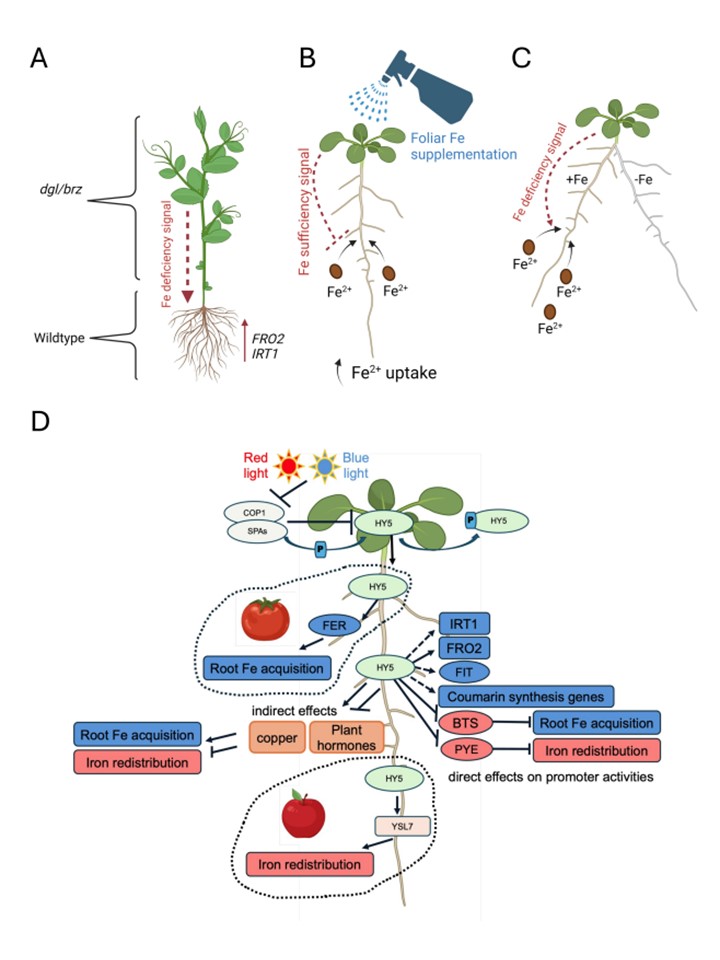
図4