鉄毒耐性に関連する候補遺伝子と有利なハプロタイプ
鉄毒耐性に関連する候補遺伝子と有利なハプロタイプ
Candidate Genes and Favorable Haplotypes Associated with Iron Toxicity Tolerance in Rice
Siyu Miao,† Jingbing Lu,† Guogen Zhang Jing Jiang Pingping Li Yukang Qian
Wensheng Wang
Int. J. Mol. Sci. 2024, 25, 6970. https://doi.org/10.3390/ijms25136970
要旨:鉄(Fe)毒性は、世界のイネ生産に悪影響を及ぼす大きな問題である。残念ながら、イネにおける鉄毒性耐性の基礎となる生理学的および遺伝学的メカニズムは比較的未知のままである。本研究では、イネ551アクセッションからなる多様なパネルを用いてゲノムワイド関連研究を実施し、鉄毒耐性に関連する遺伝的メカニズムと候補遺伝子を同定した。その結果、1、2、5、12 番染色体上に検出された 29 個の鉄 毒性耐性の量的形質座位(QTL)のうち、ハプロタイプ解析およびバイオインフォマティクス解析に より、5 個の遺伝子(qSH_Fe5、qSFW_Fe2.3、qRRL5.1、qRSFW1.1、qRSFW12)を候補遺伝子として選択した。その結果、以下の5つの遺伝子が有望な候補であることが明らかになった: LOC_Os05g40160、LOC_Os05g40180、LOC_Os12g36890、LOC_Os12g36900、LOC_Os12g36940。対照的な鉄毒耐性を示すイネ系統の生理学的特性は、イネに対する鉄毒性の悪影響を緩和するために、活性酸素種を消去する抗酸化酵素と鉄のホメオスタシスが重要であることを反映していた。今回の研究成果により、イネの鉄毒耐性の基盤となる遺伝的・生理的メカニズムが明らかになった。さらに、今後の機能解析やマーカー支援選抜による鉄毒耐性イネ品種開発のための貴重な遺伝資源を同定した。
1. はじめに
植物の必須元素である鉄(Fe)は、クロロフィル生合成、光合成、呼吸、電子輸送、酸化還元反応など、多様な生理過程に影響を与える。鉄には、第二鉄(Fe3+)と第一鉄(Fe2+)という2つの共通の原子価がある。この2つの原子価状態の間の酸化還元サイクルは、植物における鉄の取り込みと移動に不可欠であるが、活性酸素種(ROS)の生成に関連するため、鉄の毒性にも関与する。鉄の毒性を回避しながら十分な鉄を摂取するために、植物は2つの異なる戦略を進化させてきた。イネ科植物を除くすべての植物が用いる戦略Iは、プロトンを根圏に放出して土壌pHを下げるとともに、Fe3+-キレート還元酵素遺伝子の発現を誘導してFe3+をより可溶性の高いFe2+の形に還元し、これを鉄調節トランスポーター(IRT)を介して細胞内に輸送する。イネ科植物は、ムギネ酸系フィトシデロフォア(MA)の合成と土壌中への放出を伴う戦略IIを適用している。根圏のFe3+はMAとキレートを形成し、YS1(yellow stripe 1)およびYSL(yellow stripe1-like)トランスポーターを介して細胞に吸収される。イネ科植物は、ムギネ酸系フィトシデロフォア(MA)の合成と土壌中への放出を伴う戦略IIを適用している。根圏のFe3+はMAとキレートを形成し、YS1(yellow stripe 1)およびYSL(yellow stripe1-like)トランスポーターを介して細胞に吸収される。イネ(Oryza sativa L.)は戦略II植物と考えられているが、水中条件下での生育に適応しており、根圏のFe2+含量は比較的高い。そのため、イネはFe2+を吸収するメカニズムを進化させてきた。具体的には、土壌中のFe2+は主にOsIRT1とOsIRT2を介してイネ細胞内に輸送される。しかし、酸性土壌から過剰量のFe2+が吸収されると、葉細胞のFe2+濃度は有毒レベルまで上昇する。イネは、Fe濃度が300 mg L-1未満の土壌でも最適に生育する。しかし、濃度が 300 mg L-1 を超えると、イネは鉄毒性の症状を示す。重度の毒性条件下では、鉄濃度は 1000 mg L-1 にもなる。鉄毒性は、アジアや西アフリカの一部を含む世界の主要なコメ生産地域において、氷の収量を10〜90%減少させる可能性がある。イネにおける鉄毒性の特徴的な症状には、葉の "青銅色化 "と根の成長阻害がある。鉄の毒性は主に、フェントン反応を介した活性酸素とヒドロキシラジカル(-OH)の生成によるもので、その結果、細胞損傷と生理的プロセスの混乱を引き起こす。 さらに、鉄の過剰は、他のイオンの大 根への取り込みを阻害する。鉄毒性に対処するため、イネは少なくとも3つの耐性メカニズムを進化させてきた:(1) 根レベルでの鉄2+の排除とシュート問題への鉄2+ダメージの回避、(2) 過剰な鉄のアポプラストまたは液胞への貯蔵、およびプラスチドのフェリチンによる鉄の吸着、(3) 活性酸素の酵素的解毒。イネの鉄毒性耐性は、複数の遺伝子によって制御される複雑な形質である。数多くの研究により、様々な条件下でイネの鉄毒性耐性に関する多くの主効果型量的形質座位(QTL)およびエピスタティックQTLが同定されている。例えば、Liu らは MH63 と 02428 に由来する双方向性輸入系統集団を基に、イネの鉄毒性耐性に関連する 9 つの QTL を同定した。Mengらは、イネMAGIC集団を用いて鉄毒耐性に関連する9つの形質を解析し、鉄毒耐性に関連する30のQTLを検出した。さらに、イネの鉄毒性応答に寄与する遺伝子もいくつか同定された。一方、OsVIT1とOsVIT2は、過剰な亜鉛と鉄イオンを液胞に輸送し、細胞内の鉄ホメオスタシスを維持するタンパク質をコードしている。さらに、sFRO1はイネの細胞質と液胞の間の鉄ホメオスタシスの維持に重要である。最近、ゲノムワイド関連研究(GWAS)に基づく研究が、さまざまなイネ集団における鉄毒耐性に関連するQTL/遺伝子を同定するために行われている[45-51]。Matthus らは、329 のイネのアクセシ ョンを含む GWAS を実施し、鉄ストレス条件下での葉面酸化還元ホメ オスタシスの維持が鉄毒性耐性に重要であると結論づけた。Liらは、GWASと対立遺伝子相補に基づいて、シロイヌナズナの根の鉄毒性耐性の主要なQTLに関連するS-ニトロソグルタチオン還元酵素遺伝子(GSNOR)の変異体を同定した。Zhangらは、22のインディカイネ生殖質を用いてGWASを実施し、イネの鉄毒性耐性に関する14のQTLを同定した。Diopらは、有望なGWASシグナルと、高鉄ストレス応答に関与する推定候補遺伝子を同定した。イネでは鉄毒性耐性の QTL がいくつか同定されているが、Geng/Japonica(GJ)亜集団における鉄毒性耐性の遺伝的基盤や、Xian/Indica(XI)亜集団と GJ 亜集団の違いは依然として不明である。本研究では、イネの鉄毒耐性を明らかにするために、3000 Rice Genomes Project (3K-RG)の551アクセッションを用いてGWASを行った。また、代表的な鉄毒耐性アクセッションと鉄毒感受性アクセッションの鉄毒に対する生理学的応答を比較した。本研究で得られた知見は、鉄毒耐性の根底にある複雑なメカニズムを解明するものであり、今後の遺伝子の機能解明やイネの遺伝的改良に関する研究に役立つものと期待される。
3. 考察
植物では、鉄の毒性は光合成、呼吸、窒素同化を含む多様な生物学的プロセスを破壊し、最終的に細胞の損傷につながる。イネの鉄毒性耐性は、複数の遺伝子によって制御される複雑な形質である。感受性の高いイネ品種と耐性の高いイネ品種の交配から得られた遺伝子集団の解析により、鉄毒性耐性に関連する数多くのQTLが同定された。本研究では、同定された鉄毒性耐性の遺伝子座の 30%が、qSH_Fe7、qSFW_Fe2.1、qSFW_Fe3.1、qSFW_Fe12、qRSFW3、qRSH1.2、qRRL7、qRSFW1.2 などの既報の QTL [35,39,47]と同位置であった。有意なSNP、ハプロタイプ、遺伝子発現、機能的アノテーションを総合的に解析した結果、以下の5つの候補遺伝子が明らかになった。
5つの有意なQTL内に5つの有望な候補遺伝子(LOC_Os05g40160、LOC_Os05g40180、LOC_Os12g36890、LOC_Os12g36900、LOC_Os12g36940)が見つかった。特に、qSH_Fe5の最有力候補遺伝子であるLOC_Os05g40160は、NB-ARCドメインを含むタンパク質をコードしており、植物の成長と発達に不可欠なエフェクター認識とシグナル伝達に関与するシグナル伝達経路の制御に重要である。qSH_Fe5のもう一つの重要な候補遺伝子として同定されたLOC_Os05g40180(OsSTN8)は、光化学系II(PSII)コアタンパク質のリン酸化に寄与するセリン/スレオニンプロテインキナーゼをコードしている。osstn8変異体では、高照度条件下で活性酸素が蓄積し、チラコイド膜のPSII反応中心コアタンパク質が優先的に酸化される。本研究では、qRSFW12の候補遺伝子として、LOC_Os12g36890(OsCSLD4)、LOC_Os12g36900(OsDi19-7)、LOC_Os12g36940の3つを同定した。コードされるタンパク質のうち、OsCSLD4は細胞全体の多糖合成を制御し、アブシジン酸生合成を仲介するため耐塩性に重要である。OsDi19-7は干ばつ誘導19 (Di19)タンパク質ファミリーに属する。イネゲノムに存在する7つのDi19遺伝子のうち、OsDi19-4は乾燥耐性に関与している。LOC_Os12g36940はカルモジュリン結合タンパク質をコードしており、過剰な鉄レベルに応答するカルシウムを介したシグナル伝達に影響している可能性が示唆される。植物細胞において、Ca2+はセカンドメッセンジャーであり、様々な生物・生物ストレス応答に関与している。Sunらは、Ca2+とマイトジェン活性化プロテインキナーゼカスケードが、リンゴの根の鉄欠乏に対する反応に関連するシグナル伝達を制御していることを明らかにした。
鉄毒性に対する XI と GJ の亜集団の応答には明確な違いがあった。さらに、GJ 接種群、特に GJ-tmp は XI 接種群よりも鉄毒性に耐性があった(図 1E-G)。この知見は,Matthus ら による類似の研究の結果と一致する。この研究では,鉄 毒性ストレス条件下において,芳香族および温帯の GJ 接種は XI 接種よりもシュートの Fe2+ 濃度および葉のブロンズ化スコアが低いことが示された。本研究では、XI亜集団とGJ亜集団の間に共通の関連シグナルは見られなかったことから(図3B)、鉄毒性耐性の基礎となる遺伝子および/または機能的対立遺伝子の分布が、2つのイネ亜種間で大きく異なる可能性が示唆された。さらに、鉄 毒性耐性の候補遺伝子の有利なハプロタイプは、OC_Os05g40160Hap1、LOC_Os05g40180Hap1 LOC_Os12g36890Hap1、LOC_Os12g36900Hap1、LOC_Os12g36940Hap1 など、主に GJ 品種が持っていた。これらの結果を総合すると、イネの鉄毒耐性を媒介する分子メカニズムは亜種によって異なることが示唆される。毒性による酸化ストレスは、フリーラジカルの形成を誘発し、重要な生体分子(タンパク質や脂質など)に有害な影響を及ぼす可能性がある。植物組織における MDA 含量の増加は、細胞内の脂質過酸化の指標である。鉄毒性ストレス条件下では、イネの葉で MDA 産生が著しく増加し、最終的に細胞死に至った。Awasthiらは、鉄毒性ストレスにさらされると、根の脂質過酸化と細胞膜損傷が増加すると報告している。また、葉のクロロフィルとMDA含量の間に負の相関があることも報告されている。これらの先行研究と一致し、我々は、鉄毒性処理を行った試料(特に感受性 の高い品種)の MDA 含有量の有意な増加を検出した。Regonらは、重度の鉄毒性に対する耐性における活性酸素解毒の役割を提唱した。過剰な鉄レベルにさらされると、イネではSOD、カタラーゼ(CAT)、アスコルビン酸ペルオキシダーゼ、グルタチオン還元酵素(GR)、PODなどの抗酸化酵素の活性が高まる。例えば、鉄の毒性は、イネ品種ポッカリの新梢におけるCAT、POD、GR、SODの活性を著しく増加させ、抗酸化防御を高める。先行研究では、POD活性がFe2+によって誘導されることも示されている。これまでの研究でも、POD 活性は Fe2+ によって誘導されることが示されている。今回の研究では、シュートにおける POD 活性は、耐性のある系統で感受性のある系統よりも有意に高かった。したがって、鉄毒性に対する遺伝子型特異的な反応は、抗酸化酵素活性の違いに部分的に起因している可能性がある。さらに、根の Fe2+ 濃度は新梢と比較して高く、根系が鉄毒性の影響を受ける主要なイネ組織であることが示唆された。鉄毒性に耐性のある品種が、鉄イオンのホメオスタシスを維持する能力も、その耐性に寄与している可能性がある。今後の研究では、鉄毒ストレス条件下における活性酸素消去抗酸化酵素と鉄イオンのホメオスタシスを制御するメカニズムを総合的に研究する必要がある。
5. 結論
コメは世界人口の半数以上が消費しているが、一部の地域では、土壌中の過剰な鉄の蓄積がコメ栽培に悪影響を及ぼしている。この問題に対処するため、我々は現在、過剰な鉄レベルに耐えるイネ品種の分析を行っている。しかし、イネの鉄毒耐性の基礎となる生理学的・遺伝学的メカニズムは十分に理解されておらず、そのため、新しい鉄耐性イネ品種の開発が遅れている。本研究では、鉄毒耐性に関連する分子機構と候補遺伝子を明らかにするため、イネ551アクセッションからなる多様なパネルを用いてGWASを実施した。その結果、1番、2番、5番、12番染色体上に合計29個の鉄毒耐性関連QTLが検出された。これらのQTLにはqSH_Fe5、qSFW_Fe2.3、qRRL5.1、qRSFW1.1、qRSFW12が含まれ、ハプロタイプ解析とバイオインフォマティクス解析によって候補遺伝子をスクリーニングするために選択された。その結果、5つの有望な候補遺伝子が同定された(LOC_Os05g40160、LOC_Os05g40180、LOC_Os12g36890、LOC_Os12g36900、LOC_Os12g36940)。鉄毒性耐性が異なるイネ系統の生理形質から、鉄毒性によるイネへの悪影響を緩和するためには、活性酸素消去抗酸化酵素と鉄ホメオスタシスが重要であることが示唆された。この研究結果は、イネの鉄毒性耐性の基礎となる遺伝的・生理的メカニズムについての理解を深めるものであり、商業栽培イネ品種のさらなる改良に活用できる可能性がある。
以下、図1と図5の説明
図1. 鉄毒性による表現型の変化。A)地上部の高さ(SH)、(B)根の長さ(RL)、(C)シュート新鮮重(SFW)のコントロールおよび鉄毒性処理後の分布。(D) 全個体群における形質値の比の分布。(E) XIおよびGJ亜集団における形質値の比の分布。(F)XI-1A、XI-1B、XI-2、XI-3、XI-admの形質値の比率の分布。(G) GJ-adm、GJ-亜熱帯(GJ-sbtrp)、GJ-温帯(GJ-tmp)、GJ-熱帯(GJ-trp)の形質値の比の分布。鉄毒性('形質名_Fe')および対照('形質名_mock')処理については、形質値の比(R + '形質名')を示す。
図5. qRSFW12の候補遺伝子解析。(A)qRSFW12のリードSNPを囲むローカルマンハッタンプロット(上)とLDヒートマップ(下)。(B)LOC_Os12g36890のハプロタイプ解析。(C)LOC_Os12g36890のハプロタイプ間のRSFWの違い。(D)XIおよびGJ亜集団における3つのLOC_Os12g36890ハプロタイプの頻度。(E) 鉄毒性および対照条件下でのLOC_Os12g36890発現レベル。データはPlant Public RNA-Seq Databaseから得た。インディカ米品種はHachaとLachit。(F)LOC_Os12g36900のハプロタイプ解析。(G)LOC_Os12g36900のハプロタイプ間のRSFWの違い。(H)XIおよびGJ亜集団における3つのLOC_Os12g36900ハプロタイプの頻度。(I) 耐性系統(Ganjum GedおよびLalati)および感受性系統(AginsarおよびSebat)におけるLOC_Os12g36900の発現レベル。データはPlant Expression Portal Database (SRP188072)から得た。(J)LOC_Os12g36940のハプロタイプ解析。(K)LOC_Os12g36940のハプロタイプ間のRSFWの違い。(L)XIおよびGJ亜集団における3つのLOC_Os12g36940ハプロタイプの頻度。(M) 耐性系統(Ganjum GedおよびLalati)および感受性系統(AginsarおよびSebat)におけるLOC_Os12g36940発現レベル。
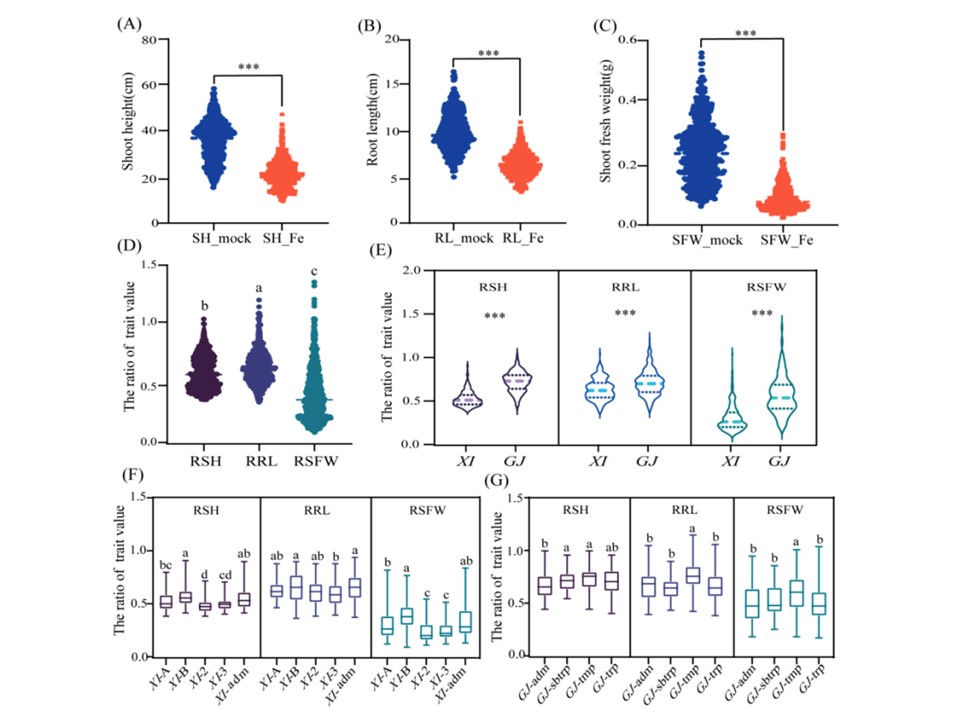
Fig1
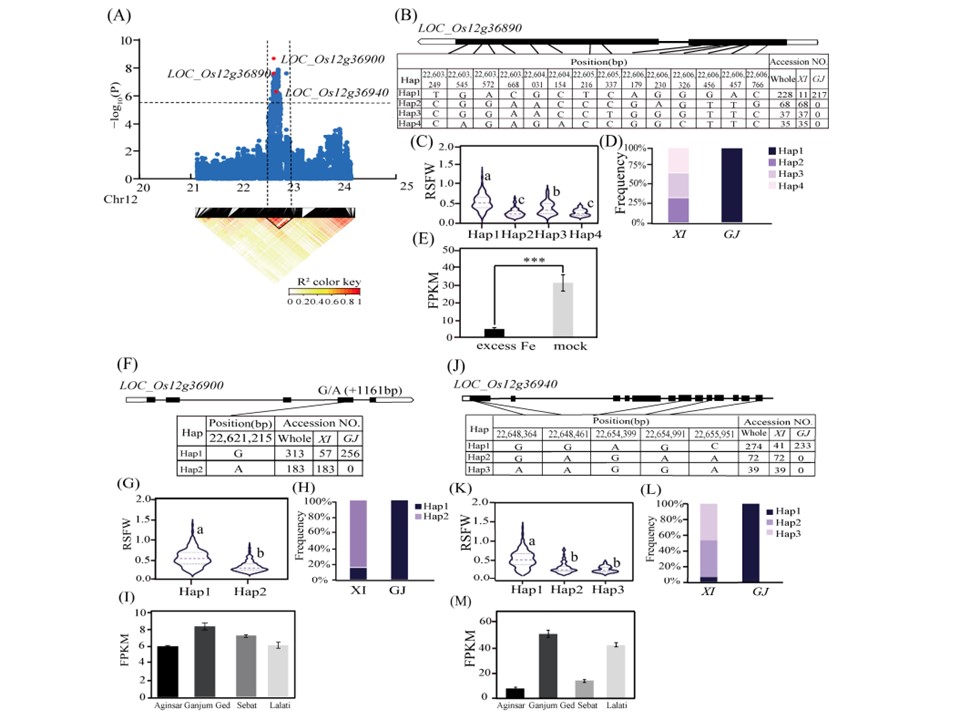
Fig5