僉儏僂儕乮Cucumis sativus L.乯偺揝寚朢墳摎妶惈壔偵偍偗傞NH4+偺僔僌僫儖揱払婡擻
僉儏僂儕乮Cucumis sativus L.乯偺揝寚朢墳摎妶惈壔偵偍偗傞NH4+偺僔僌僫儖揱払婡擻
Fatemeh Tavakoli1 - Roghieh Hajiboland1 - Dragana Bosnic2 - Predrag Bosnic3 - Miroslav Nikolic3 - Roser Tolra4 - Charlotte Poschenrieder4
Department of Plant, Cell and Molecular Biology, University of Tabriz, Tabriz, Iran 2 Institute of Molecular Genetics and Genetic Engineering, University of Belgrade, Belgrade, Serbia 3 Institute for Multidisciplinary Research, University of Belgrade, Belgrade, Serbia 4 Plant Physiology Laboratory, Bioscience Faculty, Autonomous University of Barcelona, Barcelona, Spain
庡側寢榑
NH4+ 偼丄怉暔偺娨尦宆揝寚朢斀墳傪廫暘偵婡擻偝偣傞偨傔偵昁梫偱偁傞丅
梫巪
拏慺乮N乯偼庡偵徤巁墫乮NO3-乯傑偨偼傾儞儌僯僂儉乮NH4+乯偲偟偰搚忞拞偵懚嵼偡傞丅NO3-偲NH4+偑僶儔儞僗傛偔嫙媼偝傟傞偙偲偑嵟揔側惗堢偵廳梫偱偁傞偙偲偼堦斒偵擣傔傜傟偰偄傞偑丄揝乮Fe乯妉摼偵偍偗傞偦偺廳梫惈偵偮偄偰偼廫暘偵尋媶偝傟偰偙側偐偭偨丅杮尋媶偱偼丄悈峩嵧攟偝傟偨僉儏僂儕乮Cucumis sativus L. cv. Maximus乯偵丄-Fe忦審壓偱桞堦偺拏慺尮偲偟偰NO3-傪嫙媼偟偨丅僋儘儘僔僗偑弌尰偟偨帪揰偱丄崻傑偨偼梩偵2 mM NH4Cl傪曗媼偟偨丅NH4+張棟偼丄梩偺SPAD偲HCl拪弌壜擻Fe擹搙傪憹壛偝偣偨偑丄崻偺傾億壜慪惈Fe偼尭彮偝偣偨丅崻偺堦巁壔拏慺擹搙偲FRO偺妶惈偑摨帪偵憹壛偟丄僄僠儗儞嶌梡慾奞嵻偵傛偭偰偦偺妶惈偑徚幐偟偨偙偲偐傜丄NH4+張棟怉暔偱偼Strategy-I
偺峔惉梫慺偑妶惈壔偟偨偙偲偑帵偝傟偨丅傾儞儌僯僂儉偱張棟偟偨怉暔偼丄婓敄壜梟惈偺 Fe(OH)3 偺棙梡擻偑崅偔丄H+丄 僼僃僲乕儖丄桳婡巁偺崻偐傜偺曻弌偑懡偐偭偨丅揝寚朢墳摎偺儅僗僞乕儗僊儏儗乕僞乕乮FIT乯偲偦偺壓棳堚揱巕乮AHA1丄FRO2丄IRT1乯丄偍傛傃EIN3偲STOP1偺敪尰偼丄NH4+巤梡偵傛偭偰憹壛偟偨丅帪娫揑夝愅偲僗僾儕僢僩儖乕僩僔僗僥儉偺嵦梡偵傛傝丄NH4+偑2 mM埲壓偺擹搙偱峆忢揑偵懚嵼偡傞偙偲偼丄枹抦偺僔僌僫儖傪惗惉偡傞偺偵廫暘偱偁傝丄揝寚朢徢娭楢堚揱巕偺帩懕揑側傾僢僾儗僊儏儗乕僔儑儞傪堷偒婲偙偟丄偦偺寢壥丄揝妉摼婡峔偑憹嫮偝傟傞偙偲偑帵嵈偝傟偨丅偙偺寢壥偼丄NH4+偑丄怉暔偺娨尦偵婎偯偔揝寚朢墳摎偵偍偄偰丄峀斖埻偵懚嵼偟丄偙傟傑偱偁傑傝擣幆偝傟偰偄側偐偭偨峔惉梫慺偱偁傞偙偲傪帵偟偰偄傞丅
寢榑
NO3儀乕僗偺攟抧偱摨帪偵惗堢偝偣偨怉暔偵掅擹搙偺NH4+傪揔梡偡傞偙偲偼丄掅擹搙偺NH4+乮2 mM乯偑丄塰梴揑栶妱丄H+曻弌岠壥丄偍傛傃乛傑偨偼桞堦偺拏慺尮偲偟偰巊梡偟偨応崌偺NH4+偺撆惈傪挻偊偨僔僌僫儖揱払婡擻傪帩偮偙偲傪帵偟偰偄傞丅梩偲崻偺嵶朎撪 NH4+ 儗儀儖偼丄崻偲梩偺 NH4+ 巤梡偑暘愅偟偨揝寚朢儅乕僇乕偵媦傏偡塭嬁偺掱搙偲憡娭偟偰偄側偐偭偨偙偲偐傜丄梩偲崻偺 NH4+ 巤梡偵傛偭偰惗惉偝傟丄摫娗偲巘娗傪堏摦偟丄揝寚朢偵懳偡傞憹嫮偝傟偨挿婜揑側墳摎傪曐徹偡傞壓棳偺暘巕偑婑梌偟偰偄傞偙偲偑帵嵈偝傟偨丅偦偺傛偆側僔僌僫儖偺惈幙偼晄柧偱偁傞偑丄NO/GSNO丄僄僠儗儞丄億儕傾儈儞丄傑偨偼H+乮STOP1傑偨偼NO傪夘偡傞乯偑桳椡側岓曗偱偁傞丅崻偲梩偺巤梡検偺堘偄傗丄宱帪揑側夝愅寢壥偐傜丄揝墳摎堚揱巕偺帩懕揑側傾僢僾儗僊儏儗乕僔儑儞偵偼丄2 mM傛傝偝傜偵掅擹搙偺NH4+偑峆忢揑偵懚嵼偡傞昁梫偑偁傞偙偲偑帵嵈偝傟偨丅杮尋媶偱偼丄-Fe 攟抧拞偵 Fe 尮偑懚嵼偟側偄偨傔丄梩偺 Fe 忬懺傪桳堄偵憹壛偝偣傞偙偲偼偱偒側偐偭偨偑丄搚忞惗堢怉暔偵 NH4+傪巤梡偡傞偙偲偱丄崻偺傾億僾儔僗僩偵偍偗傞 Fe 偺妉摼偲摦堳丄偍傛傃僔儏乕僩傊偺桝憲偑偐側傝夵慞偝傟傞偲婜懸偝傟傞丅偦偺寢壥 梩柺嶶晍偺曽偑崻柺嶶晍傛傝傕桪傟偨岠壥傪帵偟偨偑丄偙傟偼慜幰偑搚忞惉暘偲偺憡屳嶌梡傗旝惗暔傪夘偟偨懝幐側偟偵丄惗堢婜傪捠偠偰挿婜娫偵傢偨偭偰岠壥傪帩懕偝偣傞偙偲偑偱偒傞偐傜偱偁傞丅
埲壓丄恾偑懡偄偺偱丄傢偐傝傗偡偄恾6偲恾7偺傒傪宖嵹偟偨丅偟偐偟慡恾偺愢柧偺東栿偼丄慡晹偟偰偍偄偨丅
恾1
僉儏僂儕乮Cucumis sativus L. cv. Maximus乯偺僔儏乕僩乮a乯偲崻乮b乯偺僶僀僆儅僗偍傛傃梩偺 SPAD乮c乯丅NH4+攟抧偱揝側偟乮-Fe乯傑偨偼廫暘側揝嫙媼乮+Fe乯 偱4擔娫嵧攟偟偨屻丄CaCO3娚徴梴塼乮2.0 mM丄pH 5.8乯偱堎側傞揝張棟偲摨帪偵丄崻乮+ NH4R乯傑偨偼梩乮+ NH4L乯傪捠偟偰丄揝側偟乮-NH4+乯偍傛傃2 mM NH4+乮NH4Cl偲偟偰乯傪娷傓堎側傞 NH4+儗儀儖偱2廡娫張棟偟偨丅僨乕僞偼暯嬒亇SD (n=4)丅堎側傞暥帤偱帵偟偨朹僌儔僼娫偺嵎偼摑寁揑偵桳堄偱偁傞乮P<0.05乯丅
恾2
僉儏僂儕(Cucumis sativus L. cv. Maximus)偺庒梩拞偺HCl拪弌揝(a)偲慡揝(b)丄崻拞偺傾億僾儔僗僥傿僢僋揝(c)偲慡揝(d)偺擹搙丅怉暔偼丄-NH4+攟抧偱4擔娫Fe傪嫙媼偣偢偵惗堢偝偣偨屻丄CaCO3揧壛梴塼乮2.0 mM丄pH 5.8乯拞偱-Fe張棟偲摨帪偵丄NH4+傪崻偐傜乮+ NH4R乯丄梩偐傜乮+ NH4L乯丄傑偨偼枹張棟偺傑傑乮-NH4乯偱2廡娫張棟偟偨丅僨乕僞偼暯嬒亇SD乮n=4乯丅堎側傞暥帤偱帵偟偨朹僌儔僼娫偺嵎偼摑寁揑偵桳堄偱偁傞丅
恾 3 -NH4+ 攟抧偱 4 擔娫 Fe 傪嫙媼偣偢偵惗堢偝偣偨屻丄CaCO3 傪揧壛偟偨梴塼 (2. 0 mM) 偱 -Fe 張棟偲摨帪偵崻 (+ NH4R) (d-f) 傑偨偼梩 (+ NH4L) (g-i) 傪捠偟偰丄-NH4+ 側偟 (-NH4+) (a-c) 偍傛傃 2 mM NH4+ (as NH4Cl) 偱 4 擔娫張棟偟偨傕偺偵偍偗傞 DAF 愼怓傪梡偄偨堦巁壔拏慺 (NO) 偺専弌丅 0 mM, pH 5.8乯丅cPTIO乮NO僗僇儀儞僕儍乕乯傪僱僈僥傿僽僐儞僩儘乕儖偲偟偰梡偄偨乮c, f, i乯丅業岝帪娫偼偡傋偰摨偠乮20 ms乯丅僗働乕儖僶乕偼200 µm傪帵偡丅
恾4
僉儏僂儕(Cucumis sativus L. cv. Maximus)偺崻偵偍偗傞堦巁壔拏慺(NO)擹搙(a, c)偲揝娨尦峺慺(FRO)妶惈(b, d)偺宱帪曄壔丅Maximus)傪-NH4+攟抧偱4擔娫惗堢偝偣丄CaCO3揧壛梴塼(2.0 mM, pH 5.8)拞偱-Fe張棟偲摨帪偵NH4+傪崻偐傜(+ NH4R)丄梩偐傜(+ NH4L)丄傑偨偼柍張棟(-NH4)偱2廡娫惗堢偝偣偨丅梩柺嶶晍張棟偱偼丄-NH4 怉暔偵忲棷悈傪嶶晍偟偨丅僨乕僞偼暯嬒亇SD乮n=4乯丅堎側傞暥帤偱帵偟偨奺帪揰偺僨乕僞娫偺嵎偼丄摑寁揑偵桳堄偱偁傞乮t専掕丄P<0.05乯丅
恾5
僉儏僂儕乮Cucumis sativus L. cv. Maximus乯偺崻偺堦巁壔拏慺乮NO乯擹搙乮a乯 偲揝娨尦峺慺乮FRO乯妶惈乮b乯偺宱帪曄壔丅-NH4 攟抧偱嵧攟偟丄崻乮+ NH4R+偍傛傃+ NH4R-乯丄梩乮+ NH4L乯丄傑偨偼柍張棟乮-NH4乯偱丄2 mM NH4乮NH4Cl 偲偟偰乯偱 2 廡娫張棟偟偨丅張棟偟偨崻懁傪+ NH4R+丄枹張棟偺崻懁傪+ NH4R-偲偟偨丅僨乕僞偼暯嬒亇SD乮n=4乯丅堎側傞暥帤偱帵偝傟偨奺帪揰偺僨乕僞娫偺嵎偼摑寁揑偵桳堄偱偁傞(P<0.05)
恾 6
僉儏僂儕乮Cucumis sativus L. cv.Maximus乯偺崻偺揝娨尦峺慺乮FRO乯妶惈乮a乯丄攟抧偺 pH乮2mM偺CaCO3 傪揧壛偟丄枅擔5.8偵挷惍乯乮b乯丄梩偺 SPAD乮c乯丅Maximus乯怉暔乮d乯偺SPAD乮c乯丅-NH4+攟抧偱4擔娫Fe傪嫙媼偣偢偵惗堢偝偣偨屻丄-Fe張棟偲摨帪偵400兪M偺僠僆棸巁嬧乮STS乯偺旕懚嵼壓傑偨偼懚嵼壓偱丄NH4+傪崻偐傜乮+ NH4R乯丄梩偐傜乮+ NH4L乯丄傑偨偼枹張棟乮-NH4乯偱1廡娫張棟偟偨丅僨乕僞偼暯嬒亇SD乮n=4乯丅堎側傞暥帤偱帵偟偨朹僌儔僼娫偺嵎偼摑寁揑偵桳堄偱偁傞 (t (P<0.05)
恾 7
揝寚朢乮+Fe乯傑偨偼揝寚朢乮-Fe乯忦審壓偱嵧攟偟偨僉儏僂儕乮Cucumis sativus乯偺-NH4攟抧偵偍偗傞H+乮a乯偍傛傃僼僃僲乕儖壔崌暔乮b乯偺崻偐傜偺曻弌棪 CaCO3揧壛梴塼乮2. 0mM丄pH 5.8乯偱1廡娫張棟偟偨丅c 偺壓抜偼丄CaCO3 傪揧壛偟偨梴塼偱丄崻乮+ NH4R乯傑偨偼梩乮+ NH4L乯偐傜 NH4+ 傪嫙媼偣偢丄傑偨偼嫙媼偟偰 1 廡娫嵧攟偟偨揝寚朢怉暔傪丄50兪M 偺擄梟惈 Fe(OH)3 傪嫙媼偟偨 -NH4 梴塼乮pH 5.8乯偵 3 擔娫堏偟偨傕偺偱偁傞丅梩偺僋儘儘僼傿儖(a+b)偲僇儘僥僲僀僪(Car)擹搙(d)丅僨乕僞偼暯嬒亇SD (n=4)丅堎側傞暥帤偱帵偟偨朹僌儔僼娫偺嵎偼丄摑寁揑偵桳堄偱偁傞乮P<0.05乯丅
恾8
僉儏僂儕乮Cucumis sativus L.乯偵偍偗傞崻偺愭抂晹乮a, d, g, j乯偺桳婡巁擹搙乮兪mol g-1 FW乯偲怉暔懱慡懱偺燌弌懍搙乮nmol g-1 FW h-1乯乮b, c, e, f, h, i, k, l乯丅 僉儏僂儕傪-NH4 攟抧偱嵧攟偟丄2 mM NH4乮NH4Cl偲偟偰乯偱崻乮+ NH4R乯丄梩乮+ NH4L乯丄傑偨偼柍張棟乮-NH4乯偱張棟偟偨丅燌弌塼偼 NH4 搳梌偺 1 廡娫屻偲 2 廡娫屻偵嵦庢丒暘愅偟丄崻偺拪弌塼偼 2 廡娫偺 NH4 張棟屻偵偺傒暘愅偟偨丅僨乕僞偼暯嬒亇SD (n=4)丅堎側傞暥帤偱帵偟偨朹僌儔僼娫偺嵎偼摑寁揑偵桳堄偱偁傞乮P<0.05乯丅
恾. 9
揝寚朢乮+Fe乯傑偨偼揝寚朢乮-Fe乯忦審壓偱嵧攟偟偨僉儏僂儕乮Cucumis sativus乯偺崻偵偍偗傞 AHA1(a, b)丄FRO2(c, d)丄IRT1(e, f)偺-NH4 攟抧偱偺憡懳敪尰乮fold change乯丄 (NH4Cl)偱 1 廡娫傑偨偼 2 廡娫張棟偟丄崻(+ NH4R)傑偨偼梩(+ NH4L)傑偨偼柍張棟(-NH4)偵偟偨丅僨乕僞偼暯嬒亇SD (n=4)丅
恾. 10 僉儏僂儕乮Cucumis sativus乯偺崻傪-NH4攟抧偱揝夁忚乮+Fe乯傑偨偼揝寚朢乮-Fe乯忦審壓偱嵧攟偟丄崻乮+NH4R乯傑偨偼梩乮+NH4L乯傪2 mM NH4 (as NH4Cl)偱1廡娫傑偨偼2廡娫張棟偟偨傕偺丄傑偨偼揝張棟偲摨帪偵柍張棟乮-NH4乯偵偟偨傕偺偵偍偗傞EIN3 (a, b)丄FIT (c, d)丄STOP1 (e, f)偺憡懳敪尰乮攞悢曄壔乯丅
僨乕僞偼暯嬒亇SD (n=4)丅
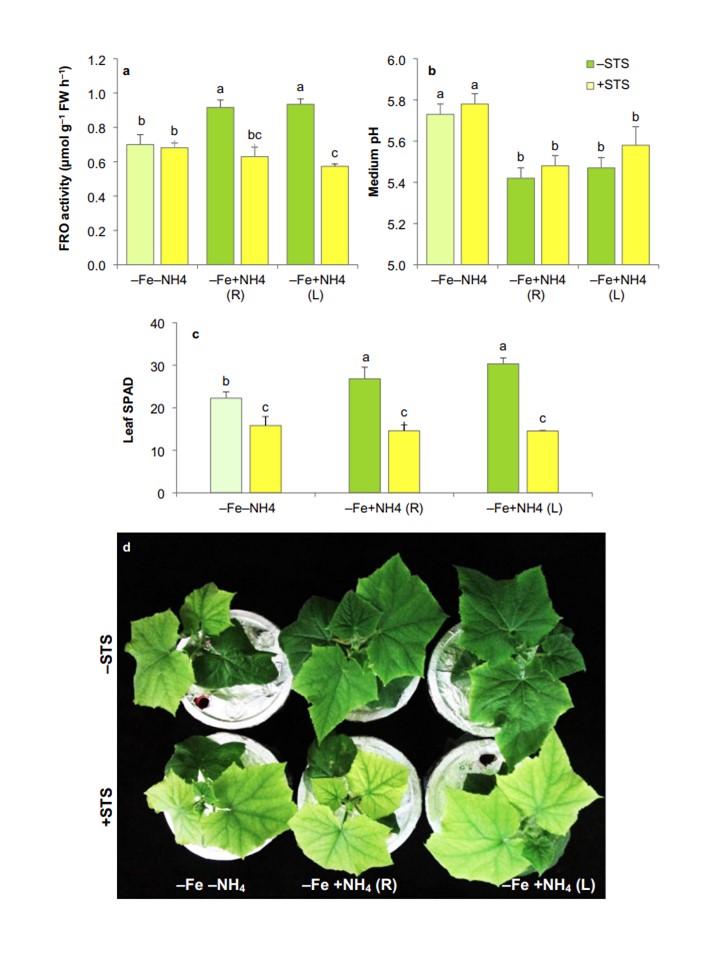
Fig.6
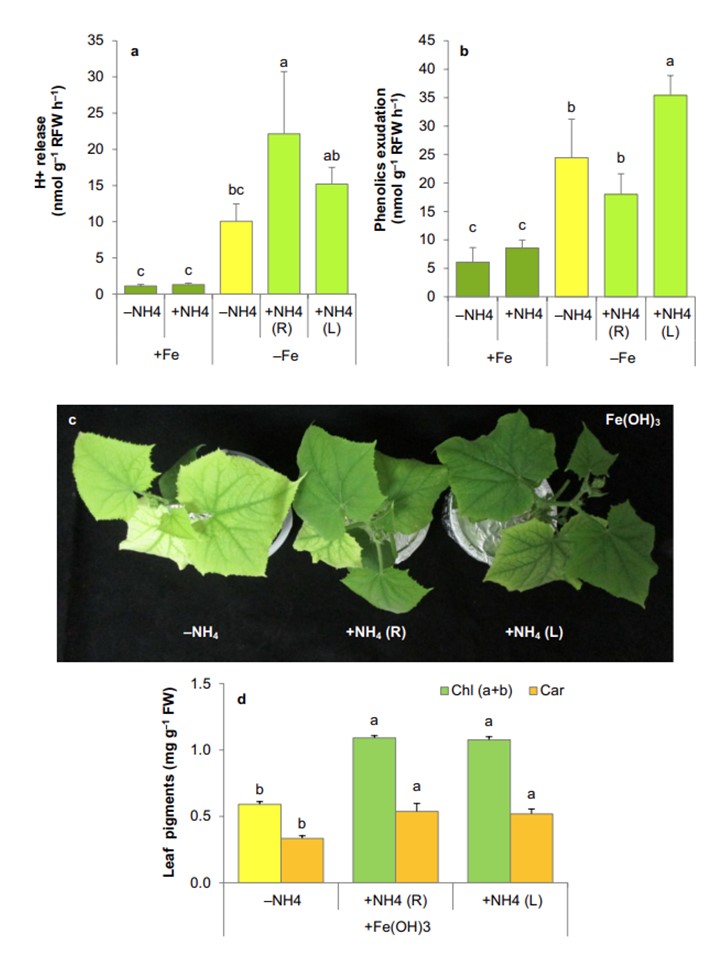
Fig.7