搤婜偵懡壥晧壸壓偱揝寚朢徢忬傪帵偡僩儅僩偺暘愅
偙偺榑暥偼僩儅僩嵧攟偺尰応偺娤嶡偐傜丄揝寚朢徢傪敪尒偟丄揝寚朢偵傛傞惗嶻惈偺掅壓偼丄愇奃幙傾儖僇儕搚忞偩偗偱側偔丄掅壏丄掅擔徠丄壥幚旍戝側偳偺僗僩儗僗忦審壓偱傕婲偙傞偙偲丄擇壙揝巤旍偵傛偭偰夞暅偡傞偙偲傪帵偟偨傕偺偱偁傞丅
偙偺愮梩導偺擾壠偵偼彫惗傕挊幰傜偲嫟偵娤嶡偟偨偙偲偑偁傝丄偦偺帪偵彫惗傕寖偟偄揝寚朢徢傪専抦偟偨婰壇偑偁傞丅偦傟傪挊幰偨偪偼暘巕儗儀儖偱偺抦尒傪摜傑偊偰棫攈側榑暥偵巇忋偘偨傛偆偩丅
丂揝寚朢偵傛傞惗嶻惈偺掅壓偼丄愇奃幙傾儖僇儕搚忞偩偗偱側偔丄掅壏丄掅擔徠丄壥幚旍戝側偳偺僗僩儗僗忦審壓偱傕婲偙傞偙偲丄擇壙揝巤旍偵傛偭偰夞暅偡傞偙偲傪帵偟偨傕偺偱偁傞丅
Analysis of tomatoes showing iron deficiency symptoms in winter under heavy fruit load
搤婜偵懡壥晧壸壓偱揝寚朢徢忬傪帵偡僩儅僩偺暘愅
Takashi Tsukamoto a , Haruhiko Inoue b , Tomoko Yokoyama a , Seiji Nagasaka c , and Yuko Ogo d,e
a Chiba Prefectural Agriculture and Forestry Research Center, Chiba, Japan; b Institute of Agrobiological Sciences, National Agriculture and Food Research Organization, Ibaraki, Japan; c Department of Life Sciences, Toyo University, Gunma, Japan; d Institute of Crop Science, National Agriculture and Food Research Organization, Ibaraki, Japan; e Institute of Vegetable and Floriculture Science, National Agriculture and Food Research Organization, Ibaraki, Japan
JOURNAL OF PLANT NUTRITION 2024, VOL. 47, NO. 10, 1650–1663 https://doi.org/10.1080/01904167.2024.2316012
梫巪
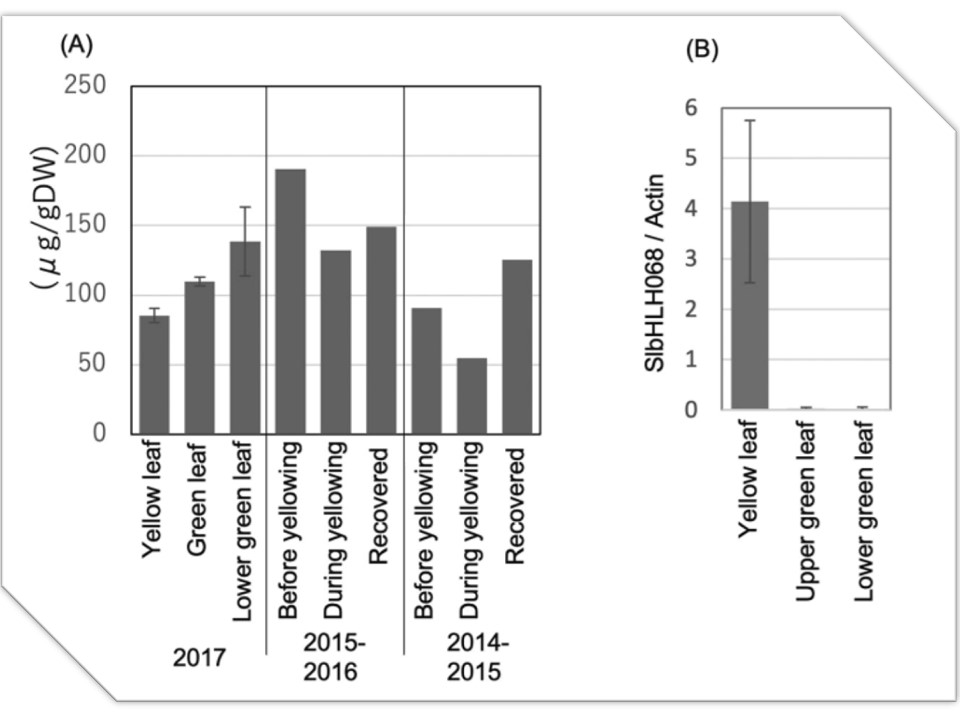
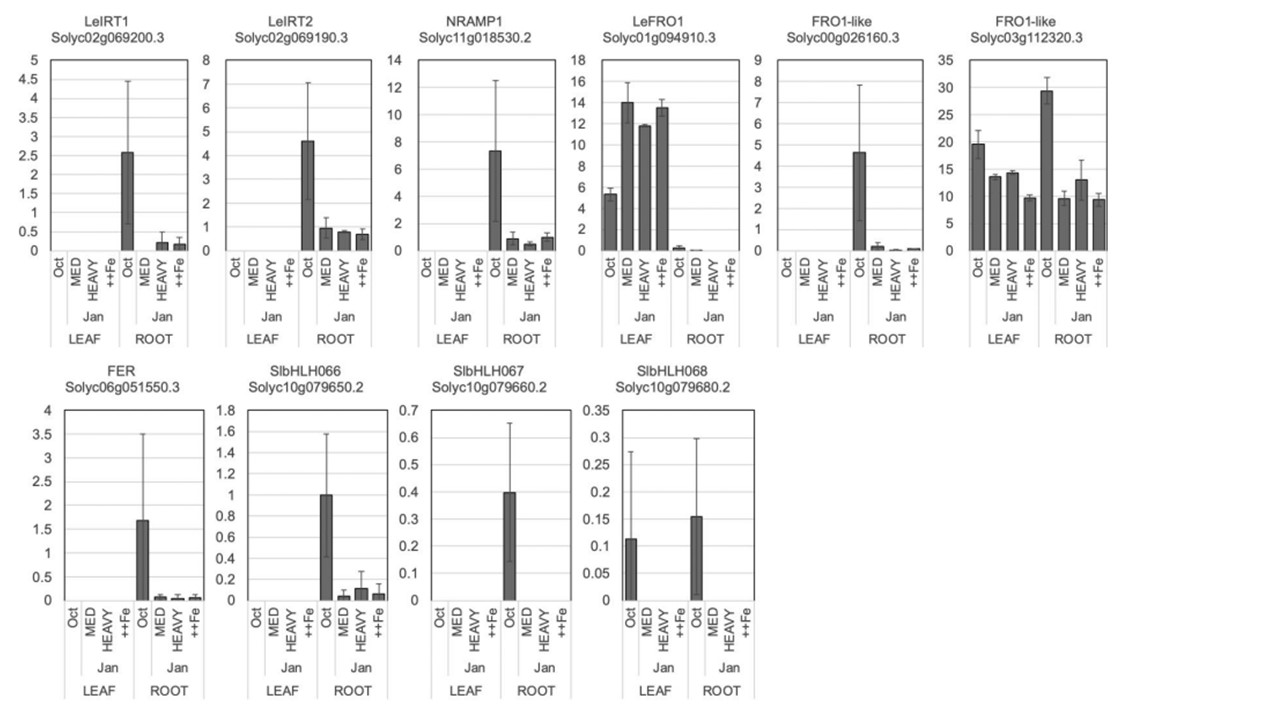
揝寚朢偵婲場偡傞偲峫偊傜傟傞嶌暔偺惗嶻惈偺掅壓偼丄愇奃幙偱側偄傾儖僇儕搚忞偱傕曬崘偝傟偰偄傞丅椺偊偽丄僩儅僩偼搤婜偵壥幚晧壸偑戝偒偄偲揝寚朢徢忬乮怴梩偺墿壔乯傪帵偡丅杮尋媶偺栚揑偼丄壥幚晧壸丄掅壏丄掅擔徠偲揝寚朢偺娭學傪挷嵏偟丄揝娷桳旍椏偵傛傞惗嶻惈岦忋偺壜擻惈傪扵傞偙偲偱偁傞丅搤婜偵揝寚朢徢忬偑敪惗偟偨擾応傪挷嵏偟偨寢壥丄揝擹搙偑掅偔丄墿壔梩偵偍偄偰揝寚朢桿敪堚揱巕偺敪尰偑忋徃偟偰偄偨丅壥幚偺廳晧壸偑揝塰梴偵媦傏偡塭嬁傪挷傋傞偨傔丄怉暔傪僩儔僗偁偨傝偺壥幚悢偑拞掱搙偺傕偺乮MED乯丄壥幚悢偑懡偄傕偺乮壥幚廳晧壸丟HEAVY乯丄壥幚悢偑懡偔揝傪捛旍偟偨傕偺乮++Fe乯偺3偮偺僌儖乕僾偵暘偗偰嵧攟偟偨丅怴梩偺SPAD梩怓抣偲搤婜偺壓梩偺廳検偼丄HEAVY亙++Fe亙MED偺弴偱偁偭偨丅偡傋偰偺僌儖乕僾偵偍偄偰丄1寧偺忋梩偺揝擹搙偼10寧偲3寧偺偦傟傛傝傕掅偐偭偨丅RNA-seq夝愅偺寢壥丄1寧偺崻偱偼揝媧廂偵娭梌偡傞堚揱巕偺敪尰偑梷惂偝傟偰偍傝丄偙傟偑搤婫偺揝寚朢偵婑梌偟偰偄傞壜擻惈偑帵偝傟偨丅偙傟傜偺寢壥偐傜丄壥幚晧壸偺戝偒偄搤婜偺僩儅僩偺惗嶻惈傪惂尷偡傞梫場偺傂偲偮偼揝偱偁傝丄揝帒嵽偵傛偭偰惗嶻惈傪夞暅偱偒傞偙偲偑帵嵈偝傟偨丅
偼偠傔偵
揝偼怉暔偵偲偭偰昁恵塰梴慺偱偁傝丄屇媧傗岝崌惉側偳條乆側惗暔妛揑僾儘僙僗偵娭梌偟偰偄傞乮Guerinot and Yi 1994乯丅悽奅偺擾抧偺30亾偼揝偑捑揳偟傗偡偄愇奃幙傾儖僇儕搚忞偱偁傞偨傔丄揝寚朢偵傛傞嶌暔偺惗嶻惈偲昳幙偺掅壓偼悽奅揑側擾嬈栤戣偱偁傞乮Marschner 1995乯丅崅摍怉暔偼丄揝傪妉摼偡傞偨傔偵2偮偺愴棯傪敪払偝偣偰偒偨乮Kobayashi and Nishizawa 2012乯丅僀僱壢怉暔偼丄揝僉儗乕僞乕偱偁傞儉僊僱巁傪娷傓愴棯II傪梡偄傞丅旕僀僱壢怉暔偼丄崻寳偵僾儘僩儞傪曻弌偟丄揝傪傛傝壜梟惈偺崅偄戞堦揝偵娨尦偟偰媧廂偡傞愴棯I傪梡偄傞丅愴棯I傪梡偄傞僔儘僀僰僫僘僫偱偼丄嵶朎枌H+-ATPase偑僾儘僩儞傪崻寳偵曻弌偟丄揝僉儗乕僩娨尦峺慺2乮FRO2乯偑揝傪戞堦揝偵娨尦偟丄IRON REGULATED TRANSPORTER 1乮IRT1; Robinson et al.乯 揝媧廂偵娭梌偡傞堚揱巕傪惂屼偡傞揮幨場巕傕摨掕偝傟偰偄傞丅FER條揝寚朢桿摫揮幨場巕(FIT)偼墫婎惈/僿儕僢僋僗儖乕僾僿儕僢僋僗(bHLH)僞儞僷僋幙偱偁傝丄懠偺bHLH僞儞僷僋幙AtbHLH38丄AtbHLH39丄AtbHLH100丄AtbHLH101)偲憡屳嶌梡偟丄A. thaliana偺FRO2丄IRT1丄IRT2丄僯僐僠傾僫儈儞僔儞僞乕僛1(NAS1)丄NAS2側偳偺壓棳堚揱巕偺敪尰傪挷愡偡傞(Colangelo and Guerinot 2004; Jakoby et al. 2004; Schwarz and Bauer 2020; Wang et al. 2013; Yuan et al. 2005, 2008)丅僩儅僩偱偼丄揝塰梴偵娭楢偡傞偄偔偮偐偺堚揱巕偑摨掕偝傟丄偦偺婡擻偑夝柧偝傟偰偄傞丅LeIRT1偲LeIRT2偼丄僔儘僀僰僫僘僫偺IRT1偲傾儈僲巁攝楍偺憡摨惈偑嵟傕崅偄ZIP僩儔儞僗億乕僞乕偱偁傝丄椉幰偲傕揝偺庢傝崬傒妶惈傪帵偡乮Bereczky傜 2003; Eckhardt, Mas Marques, and Buckhout 2001乯丅LeNRAMP1偼堐娗懇偵敪尰偟丄揝僩儔儞僗儘働乕僔儑儞偵娭梌偟偰偄傞偲峫偊傜傟偰偄傞 (Bereczky et al. 2003)丅LeFRO1偼揝偺庢傝崬傒偵娭梌偡傞庡梫側揝僉儗乕僩娨尦峺慺偱偁傞 (Li, Cheng, and Ling 2004)丅FER偼FIT偺僩儅僩僆儖僜儘僌偱偁傝丄LeIRT1丄LeFRO1丄LeNRAMP1偺敪尰傪惂屼偟偰偄傞(Bereczky et al. 2003; Brumbarova and Bauer 2005; Ling et al. 2002)丅SlbHLH066丄067丄068偼僔儘僀僰僫僘僫偺bHLH揮幨場巕AtbHLH38丄AtbHLH39丄AtbHLH100丄AtbHLH101偺儂儌儘僌偱偁傞丅SlbHLH068偼FER偲憡屳嶌梡偟偰LeFRO1偺敪尰傪惂屼偟偰偄傞乮Du et al.乯 擔杮偺傛偆側巁惈搚忞偺抧堟偱偼丄搚忞拞偺揝偑梕堈偵壜梟壔偟嶌暔偵棙梡偱偒傞偨傔丄嶌暔偵偍偗傞揝寚朢偼偁傑傝拲栚偝傟偰偙側偐偭偨丅偟偐偟丄嬤擭丄旕僇儖僔僂儉丒傾儖僇儕搚忞偵偍偄偰傕丄揝寚朢偑尨場偲巚傢傟傞嶌暔偺惗嶻惈傗昳幙偺掅壓偑曬崘偝傟偰偄傞乮Suzuki, Matsuyama, and Kikuchi 2018乯丅戙昞揑側椺偲偟偰丄僩儅僩偱偼搤婜偵壥幚晧壸偑戝偒偔側傞偲揝寚朢徢條徢忬乮怴梩偺梩柆娫僋儘儘僔僗乯偑尰傟傞丅梩柆娫僋儘儘僔僗偼丄悈峩嵧攟偲搚忞嵧攟偺椉曽偱丄偝傑偞傑側昳庬丄儈僯僩儅僩傗戝嬍僩儅僩偱敪惗偟偨丅擾壠偑擇壙揝傪岠棪傛偔嫙媼偡傞揝帒嵽乮揝椡傾僋傾丟徏嶳傜2008乯傪巤梡偡傞偲丄梩柆娫僋儘儘僔僗偲廂検掅壓偑夞暅偡傞孹岦偑傒傜傟偨乮楅栘丒徏嶳丒媏抮2018乯丅嬤擭丄掅壏丒掅擔徠僗僩儗僗壓偱崻偺揝娨尦峺慺妶惈偑掅壓偡傞偙偲偑敾柧偟偰偄傞乮Suzuki, Matsuyama, and Kikuchi 2018乯丅偙傟傜偺帠椺偐傜丄揝偺媧廂偑掅壓偟偨偨傔偵僩儅僩偺廂検偑尭彮偟偨壜擻惈偑峫偊傜傟傞丅杮尋媶偱偼丄僩儅僩偱揝寚朢徢忬偑娤嶡偝傟偨愮梩導偺擾応傪挷嵏偟偨丅師偵丄幚尡壏幒偱掅壏丒掅擔徠僗僩儗僗壓丄壥幚晧壸偺堎側傞僩儅僩傪挿婜嵧攟偟丄揝偺媧廂丒暘晍偲堚揱巕敪尰傪夝愅偟偨丅捠忢偺揝寚朢偲偺斾妑偺偨傔丄奐壴婜傑偱揝傪娷傑側偄悈峩嵧攟傪峴偭偨僩儅僩偺崻偲梩偵偮偄偰RNA-seq傪峴偭偨偑丄偙傟偼僩儅僩偺抧忋晹偲奐壴屻偺揝寚朢偵偍偗傞堚揱巕敪尰偵娭偡傞尋媶偑傎偲傫偳側偐偭偨偨傔偱偁傞丅
寢榑
愮梩導偺擾応偱娤嶡偝傟偨墿壔偟偨僩儅僩偺梩偼丄揝娷検偑掅偔丄揝寚朢桿摫惈堚揱巕偺敪尰偑憹壛偟丄揝寚朢偵嬤偄忬懺偱偁偭偨丅壥幚晧壸偺堎側傞僩儅僩偺挿婜嵧攟幚尡偱偼丄1寧偵偡傋偰偺僌儖乕僾偱忋梩偺揝擹搙偑掅壓偟偨丅偙傟偼丄揮幨場巕FER偍傛傃SlbHLH066, 067, 068偺僟僂儞儗僊儏儗乕僔儑儞偵懕偒丄庡梫側揝桝憲懱LeIRT1, LeIRT2偍傛傃LeNRAMP1偺僟僂儞儗僊儏儗乕僔儑儞偵傛偭偰堷偒婲偙偝傟偨偲峫偊傜傟傞丅搤偺SPAD抣偲梩廳偺掅壓偼丄HEAVY < ++Fe < MED偺弴偱偁偭偨丅偙傟傜偺寢壥偼丄掅壏傗掅擔徠偵傛偭偰揝偺媧廂丒棙梡岠棪偑掅壓偟丄壥幚偺廳愊偑戝偒偔側傞偲怉暔偺惗嶻惈偑掅壓偡傞孹岦偑偁傝丄擇壙揝偺巤旍偵傛偭偰夞暅偡傞偙偲傪帵偟偰偄傞丅偙偺偙偲偼丄掅壏丄掅擔徠丄懡壥晧壸側偳偺忦審壓偱偼丄揝偑惗嶻惈傪掅壓偝偣傞惂尷場巕偺堦偮偱偁傝丄擇壙揝偺巤旍偵傛偭偰惗嶻惈偑夞暅偡傞偙偲傪帵嵈偟偰偄傞丅杮尋媶偱偼丄揝寚朢偵傛傞惗嶻惈偺掅壓偼丄愇奃幙傾儖僇儕搚忞偩偗偱側偔丄掅壏丄掅擔徠丄壥幚旍戝側偳偺僗僩儗僗忦審壓偱傕婲偙傞偙偲丄擇壙揝巤旍偵傛偭偰夞暅偡傞偙偲傪帵偟偨丅
乮埲壓恾偺愢柧乯
恾2. 愮梩導偺擾応偵偍偗傞僩儅僩梩偺僋儘儘僔僗丅(A乯僩儅僩梩偺梩柆娫僋儘儘僔僗丅
恾3. 愮梩導偺擾応偵偍偗傞墿壔偟偨梩偺暘愅丅(A乯僩儅僩偺梩偺揝偺擹搙丅墿壔偟偨2017擭偵丄墿壔梩丄墿壔梩偲摨偠崅偝偺椢梩丄墿壔梩偲摨偠姅偺壓晹偺椢梩傪廂妌偟偨丅僄儔乕僶乕偼昗弨曃嵎乮n= 3乯丅2014乣2016擭偼丄墿壔慜丄墿壔拞丄墿壔偐傜夞暅偟偨梩傪廂妌偟偨丅惗暔妛揑暋惢側偟丅(B乯2017擭偵廂妌偟偨梩偵偍偗傞SlbHLH068偺儕傾儖僞僀儉PCR丅僄儔乕僶乕偼昗弨曃嵎乮n = 3乯丅
恾5. 堎側傞壥幚晧壸偱偺挿婜嵧攟偵偍偗傞丄揝偺媧廂偲堏摦偵娭梌偡傞堚揱巕偺RNA-seq偺100枩儅僢僾儕乕僪偁偨傝偺僄僋僜儞1僉儘墫婎偁偨傝偺抐曅悢乮FPKM乯丅僄儔乕僶乕偼昗弨曃嵎乮n = 3乯丅堚揱巕柤偺壓偺堚揱巕嵗斣崋偼ITAG3.0僎僲儉偵傛傞丅奺孮偺娫堷偒偵傛傞壥幚悢偺挷惍偼10寧埲崀偵峴偭偨丗10寧偺壥幚晧壸偼慡孮偱摨偠丄1寧偺壥幚晧壸偼奺孮偱堎側傞丅

恾2
恾3
恾5