水芭蕉の熱発生スパディックスにおける遺伝子発現と代謝物レベルの収束
この論文の著者の伊藤菊克岩手大学教授(現農学部長)ムギネ酸の発見者である故高城成一先生の直弟子です。
Ito K, Ito T, Onda Y, Uemura M. Temperature-triggered periodical thermogenic oscillations in skunk cabbage (Symplocarpus foetidus). Plant Cell Physiol. 2004:45(3):257-264.
の論文で「ざぜん草」の”温度依存性発熱リズム”を発見して発表して以来20年間にわたって、この研究を続けてこられました。高城成一先生の精神影響を受け継がれてきたものと思われます。
Gene expression and metabolite levels converge in the thermogenic spadix of skunk cabbage
水芭蕉の熱発生スパディックスにおける遺伝子発現と代謝物レベルの収束
谷本はるか、 梅川 結衣、 高橋英之、 後藤康太、伊藤菊克
1 United Graduate School of Agricultural Science, Iwate University, Morioka, Iwate 020-8550, Japan 2 Department of Planning and General Affairs, Akita Research Institute of Food and Brewing, Araya-machi, Akita 010-1623, Japan 3 Department of Agriculture, School of Agriculture, Tokai University, Kumamoto 862-8652, Japan 4 Faculty of Agriculture, Iwate University, Morioka, Iwate 020-8550, Japan
PLANT PHYSIOLOGY 2024: 195: 1561-1585
(要旨)
水芭蕉(Symplocarpus renifolius)の花序(穂軸)は強い熱発生を示し、周囲温度が氷点下になっても23℃前後で温度を調節できる。水芭蕉の発生過程で制御される熱発生と体温調節の基礎となるメカニズムを解明するため、われわれは穂軸の発生の3つの段階にわたって、トランスクリプトームとメタボロームの包括的解析を行った。RNA-seq解析の結果、セレン結合タンパク質1/メタンチオールオキシダーゼ(SBP1/MTO)の発現が、体温を上昇させる花托で最も高かった。特に、alternative oxidase (AOX)の発現は、プレ発熱期から発熱期まで一貫して高かった。メタボローム解析から、ヌクレオチドレベルの変化が、水芭蕉の発育制御された組織特異的な熱発生に対応していることが示された。また、SBP1/MTOの産物である硫化水素は、チトクロームcオキシダーゼ(COX)を介するミトコンドリア呼吸を阻害するが、AOXを介する呼吸は比較的影響を受けないことも明らかになった。特に低温では、COXを介した呼吸に対する硫化水素の阻害効果が増大し、AOXを介した呼吸が優勢になる。最後に、棘突起の発達を通して遺伝子と代謝物の調節に差があるにもかかわらず、熱発生時には遺伝子発現と代謝物蓄積パターンの収束が観察された。この同期性は、発生制御された熱発生において重要な役割を果たしている可能性がある。さらに、このような収束は 水芭蕉の体温調節の確かな分子基盤を提供する可能性がある。
(はじめに)
特定の器官や組織における内因性の代謝熱産生プロセスである熱発生は、様々な被子植物種で観察される生理現象である。植物の熱発生は、しばしば熱発生器官からの匂いや香りの放出と関連している。注目すべきは、このメカニズムが繁殖期に花粉媒介者を引き寄せると同時に、訪花昆虫のエネルギー節約に有利な暖かい生息環境を作り出すことである。タイタンアルム(Amorphophallus titanum)、デッドホースアルム(Helicodiceros muscivorus)、クレタアルム(Arum concinnatum)など多くの植物がある、 などのキク科の植物や、ハス科の神聖ハス(Nelumbo nucifera)やアメリカハス(Nelumbo lutea)などが体温上昇を示すが、その程度や時期は種によって異なる。水芭蕉(Symplocarpus renifolius)は、周囲が氷点下になることが多い晩冬から早春にかけて開花する熱発生種である。穂状花序と呼ばれる花序は、寒い環境下で体内温度を上昇させることができる。S.renifoliusの穂軸は小葉と髄からなる。われわれが過去に行ったスパディスの発熱段階の熱分析から、胞子の外側に位置する小花は、内側の髄組織に比べて高い温度を維持することが明らかになった。従って、熱発生は主に小花で起こり、果柄はこのプロセスに直接は寄与しないことが示唆された。胞子嚢の発達初期に相当する再発熱期には、器官の構造はまだ未熟で、目に見える突起もない。この段階は、その後の生理的変化の基礎を作るもので、活動性はない。その後、穎果は雌性期に移行し、熱発生が大幅に増加し、体温調節が始まる。この段階は、その後の生理的変化の下地を作る段階であり、活動を伴わない。その後、穎果は雌性期に移行し、熱発生の大幅な増大と体温調節の開始を特徴とする。この段階では、小花は成熟し、雌しべとまだ未熟な雄しべ、そして維管束を含む髄を伴う。その後、雄しべは、雌性期と雄性期をつなぐ一時的な期間である両性期を経て、後発温期とも呼ばれる雄性期に移行する。雄性期には、雄しべは消滅し、花粉が胞子嚢の表面に現れ、熱発生はかなり減少する。レニフォリウスの距に見られるこの発育パターンは、二性交配の一例であり、同じ植物が一つの開花期間中に異なる時期に異なる性相を経る。S. renifoliusは二卵性であり、自家不和合性であり、繁殖には他の個体からの花粉を必要とするにもかかわらず、他の熱発生植物に比べ、花粉媒介者の誘引のための熱発生への依存度は低い。興味深いことに、「水芭蕉 」という通称は、北米の親戚である
Symplocarpus foetidusと共通する「スカンク・キャベツ」という通称があるにもかかわらず、どちらの種も熱発生時にスカンクに典型的に見られるような強く不快な臭いを発しない。この観察結果は、S. renifoliusのスパディスの3つの発生段階(発熱期を含む)を通して、ジメチルスルフィド(DMS)、ジメチルジスルフィド(DMDS)、ジメチルトリスルフィド(DMTS)などの臭気化合物を含む花の揮発性物質の組成に実質的な変化が観察されなかったことを報告した以前の研究結果からも裏付けられた。さらに、Calliphoridae(クロバエ科)やDead-horse arumの場合とは異なり、凍てつくような寒冷環境では、S. renifolius spadicesから放出される化合物に昆虫が引き寄せられるという証拠はほとんどない。しかし生理学的研究により、レニフォリウスでは花粉の発芽と花粉管の伸長において、調節された温度(23℃)が重要な役割を果たしていることが明らかになった。褐色脂肪組織(BAT)では、ミトコンドリア呼吸が細胞熱産生の主要な原動力であることが認識されている。熱産生は、基質の酸化から生じる熱効果を最大化するように設計されたいくつかの生化学的戦略によって増強することができる。(ii)ミトコンドリアの内膜を横切るプロトンの同時移動とは無関係に電子伝達連鎖を進行させる; (iii)プロトンリークと呼ばれるプロセスで、ATP合成を伴わない場合にミトコンドリア内膜を横切るプロトン電気化学的勾配の消滅を可能にする。哺乳類のBAT細胞では、主要な発熱エフェクターは、プロトンリーク動態を促進する重要な役割を果たすアンカップリングタンパク質1(UCP1)である。植物の熱発生は、ミトコンドリア密度を含むミトコンドリア呼吸活動や、ATP合成効率を低下させる生化学的変数によっても影響を受ける。これには主に、ミトコンドリアの電子輸送鎖におけるプロトン勾配の確立が関わっている。このプロセスにおける重要な構成要素は、ミトコンドリアの代替オキシダーゼ(AOX)であり、直接のプロトン輸送をバイパスして、ユビキノールから酸素への電子の直接移動を可能にする。このメカニズムは、電子のスリップやリークというカテゴリーと一致する。同様に、ロテノン非感受性NAD(P)Hデヒドロゲナーゼ、内部代替NAD(P)H:ユビキノン酸化還元酵素(NDA)および外部代替NAD(P)H:ユビキノン酸化還元酵素(NDB)は、植物のミトコンドリア内膜の両側に位置し、プロトンポンプに頼らずに複合体Iのバイパスとして機能する。S. renifoliusでは、AOX転写産物およびAOXタンパク質は、熱産生スパディックスにおいて著しく豊富である。さらに、AOX転写産物は小花、特に花弁と雌しべに特異的に見られる。花弁と雌しべでAOXが組織特異的に発現していることから、S. renifoliusの発育制御された熱発生において、これらの組織が重要な役割を果たしている可能性が高い。
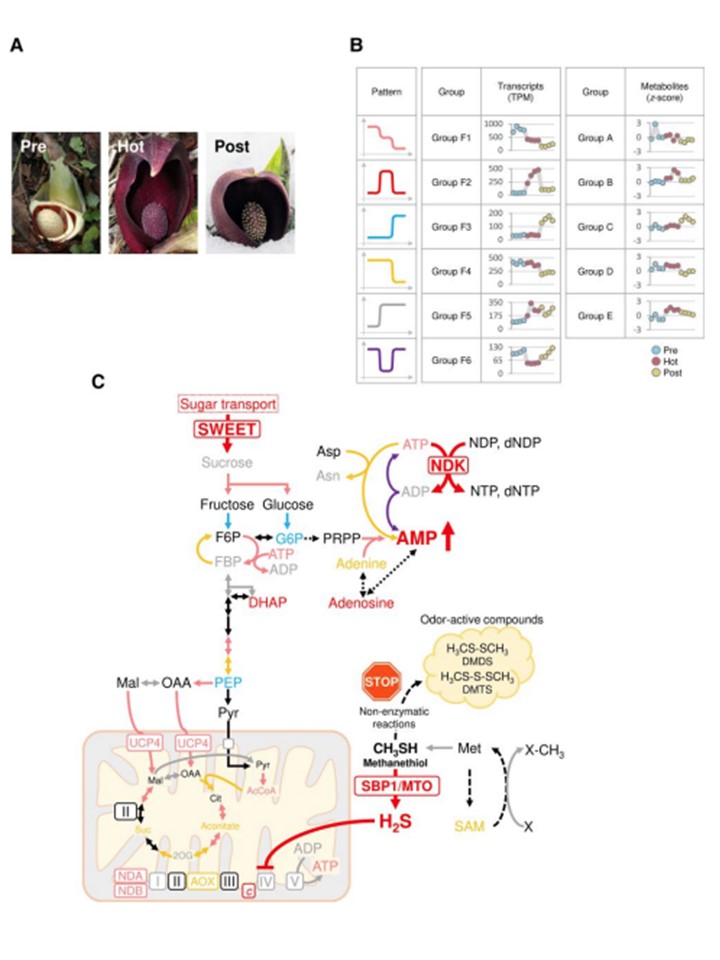
(図9)
S. renifoliusの発熱性スパディスの小花における遺伝子発現と代謝物蓄積パターンの収束と統一。A) S. renifoliusの発熱前(Pre)、発熱期(Hot)、発熱後(Post)の代表的な画像。プレステージでは胞子が取り除かれ、棘が見える。B) 異なるステージにおける小花の遺伝子発現と代謝産物蓄積のパターン。パネル中の記号は、各グループにおける明確な遺伝子発現/代謝物蓄積パターンを表す。表示されたグループは、トランスクリプトーム解析(図3B)およびメタボローム解析(図5F)に対応する。C) 小花の発生過程における遺伝子発現と代謝経路の変化の概略。パネル(B)に異なる記号で示したように、発生過程における遺伝子発現の明確なパターンは、発熱期でほぼ最大に融合する。発熱期における主要なシフトには、中心的な炭素代謝とAOXを含むミトコンドリア呼吸鎖成分のアップレギュレーションが含まれる。SWEETは解糖系経路を刺激する。ATP消費の増加は、NDKが駆動するヌクレオチド相互変換とアスパラギン合成酵素の発現によって媒介される。AMPの上昇は異化過程を刺激し、代謝熱産生の急増を引き起こす。SBP1/MTOはメタンチオールを分解して硫化水素を産生し、COX活性を抑制して代謝フラックスをAOX呼吸経路に向かわせ、代謝熱産生に寄与する。メタンチオール分解へのシフトと悪臭化合物(DMDS、DMTS)の産生の減少により、S. renifoliusの無傷の穂軸から放出される悪臭は大幅に減少した。SBP1/MTO、セレン結合タンパク質1/メタンチオールオキシダーゼ;AOX、代替オキシダーゼ;COX、シトクロムcオキシダーゼ;NDA、内部代替NAD(P)H:ユビキノンオキシドレダクターゼ;NDB、外部代替NAD(P)H:ユビキノンオキシドレダクターゼ;NDK、ヌクレオシド二リン酸キナーゼ;SWEET、糖は最終的に輸出されるトランスポーター;DMDS、ジメチルジスルフィド; DMTS、ジメチルトリスルフィド;AcCOA、アセチル-CoA;ADP、アデノシン二リン酸;AMP、アデノシン一リン酸;Asn、アスパラギン;Asp、アスパラギン酸;ATP、アデノシン三リン酸;Cit、クエン酸;dNDP、デオキシヌクレオシド二リン酸;dNTP、デオキシヌクレオシド三リン酸;DHAP、ジヒドロキシアセトンリン酸; F6P、フルクトース6-リン酸;FBP、フルクトース1,6-ビスホスフェート;G6P、グルコース6-リン酸;Mal、リンゴ酸;NDP、ヌクレオシド二リン酸;NTP、ヌクレオシド三リン酸; OAA、オキサロ酢酸;PEP、ホスホエノールピルビン酸;PRPP、5-ホスホリボシル1-ピロリン酸;SAM、S-アデノシルメチオニン;Suc、コハク酸;2OG、2-オキソグルタル酸。