IMA1偺嬻娫揑側惂屼偼MAMP傪擣幆偡傞偙偲偵傛傝崻偵傛傞揝妉摼傪惂尷偡傞
IMA1偺嬻娫揑側惂屼偼MAMP傪擣幆偡傞偙偲偵傛傝崻偵傛傞揝妉摼傪惂尷偡傞
Spatial IMA1 regulation restricts root iron acquisition on MAMP perception
Nature
https://doi.org/10.1038/s41586-023-06891-y
Received: 14 October 2022
Accepted: 22 November 2023
Published online: 10 January 2024
奣梫
揝偼廻庡偲旝惗暔偺憡屳嶌梡偵偍偄偰廳梫偱偁傞丅姶愼帪偵廻庡偑棙梡壜擻側揝傪惂尷偡傞偙偲偼丄塰梴柶塽偲屇偽傟傞廳梫側杊屼愴棯偱偁傞丅偟偐偟丄挵娗忋旂傗怉暔偺崻偺昞旂偺傛偆偵丄揝偺惗暔妛揑棙梡擻偵桳棙側娐嫬傪嶌傝弌偡丄奜晹偵柺偟偨媧廂惈偺慻怐偵偲偭偰偼丄偙傟偼擄栤偱偁傞丅椺偊偽丄怉暔偺崻偼庡偵搚忞偐傜揝傪媧廂偟丄揝偑晄懌偡傞偲丄崻寳偺巁惈壔傗揝僉儗乕僩嵻偺暘斿側偳偺儊僇僯僘儉偵傛偭偰揝偺棙梡棪傪崅傔傞丅偟偐偟丄揝偺惗暔妛揑棙梡擻偺忋徃偼丄怉暔偺寬峃傪嫼偐偡僶僋僥儕傾偺憹怋偵偲偭偰傕桳塿偱偁傞丅偙偙偱変乆偼丄僔儘僀僰僫僘僫偵偍偄偰丄僼儔僕僃儕儞側偳偺旝惗暔娭楢暘巕僷僞乕儞偑丄慡恎偺揝寚朢僔僌僫儖儁僾僠僪偱偁傞傾僀傾儞儅儞1乮IMA1乯偺嬊強揑側暘夝傪捠偠偰丄崻偺揝妉摼梷惂偵偮側偑傞偙偲傪曬崘偡傞丅
偙偺斀墳偼丄嵶嬠偑崻慻怐偵怤擖偟偨偲偒偵傕桿敪偝傟傞偑丄嵶嬠偑崻偺奜昞柺偵惗懅偟偰偄傞偲偒偵偼桿敪偝傟側偄丅IMA1帺恎偼丄崻偲僔儏乕僩偵偍偗傞柶塽傪挷愡偡傞栶妱傪帩偪丄崻偺僐儘僯乕宍惉儗儀儖偲嵶嬠惈梩柺昦尨懱偵懳偡傞掞峈惈偵塭嬁傪梌偊傞丅変乆偺敪尒偼丄揝偺惗暔妛揑棙梡擻偲庢傝崬傒丄偍傛傃柶塽墳摎偵塭嬁傪梌偊傞塰梴柶塽偺揔墳揑暘巕儊僇僯僘儉傪柧傜偐偵偟偨丅
峫嶡
変乆偺寢壥偼丄flg22 MAMP偑挿婜娫懚嵼偡傞偲丄A. thaliana偵偍偗傞揝寚朢墳摎偺庡梫側峔惉梫慺偑徚幐偡傞偙偲傪柧傜偐偵偟偨丅
惗暔妛揑偵棙梡壜擻側揝傪崻偵桝憲偟丄妘棧偡傞偙偲偱丄愽嵼揑側昦尨懱偐傜揝傪扗偆偙偲偑偱偒傞偺偵丄偙傟偼晄壜夝偵巚偊傞丅偟偐偟丄偙偺傛偆側墳摎偼丄惗暔妛揑偵棙梡壜擻側揝偑娐嫬拞偵朢偟偄応崌偵偺傒婲偙傞乮偦偆偱側偗傟偽丄怉暔偼揝寚朢偵側傜側偄乯偺偱丄揝寢崌壔崌暔傗僾儘僩儞傪燌弌偝偣傞偙偲偵傛偭偰揝傪惗暔妛揑偵棙梡壜擻偵偟懕偗傞偙偲偼丄昦尨懱偵傕棙塿傪傕偨傜偡偐傕偟傟側偄丅
埲忋偺偙偲偐傜丄IMA1傪夘偟偨揝寚朢墳摎偲flg22傪夘偟偨杊屼墳摎偲偺娫偺漢峈婡擻偼丄愽嵼揑側昦尨懱偵偲偭偰揝偑惗暔妛揑偵棙梡壜擻偵側傞偙偲傪旔偗丄怉暔偺杊屼墳摎傪懝側傢側偄偨傔偵廳梫偱偁傞壜擻惈偑帵嵈偝傟偨丅変乆偺敪尒偼傑偨丄揝寚朢墳摎偺挷愡偼丄扨偵嵶嬠偑懚嵼偡傞偙偲偵傛偭偰堷偒婲偙偝傟傞峔惉揑側墳摎偱偼側偔丄嫼埿傪帵偡崌恾乮椺偊偽丄崅儗儀儖偺flg22偺帩懕揑側懚嵼傗慻怐懝彎乯偺懚嵼偵墳偠偰堷偒婲偙偝傟傞傕偺偱偁傞偙偲傪帵偟偰偄傞丅堦斒偵巁惈乮娨尦惈乯忦審偲僋儅儕儞偼崻寳偱偺揝梟夝惈傪懀恑偡傞偨傔丄揝偑惂尷偝傟偨忦審壓偱寬慡側崻寳傪堐帩偡傞偵偼丄偙偺傛偆側偒傔嵶偐側挷愡偑廳梫偱偁傞偲巚傢傟傞丅揝偺妉摼傪掆巭偡傞偙偲偼丄桳奞側嵶嬠偑揝偵梕堈偵傾僋僙僗偱偒傞傛偆偵偡傞偙偲傪旔偗傞曽朄偐傕偟傟側偄偑丄摨帪偵丄怉暔帺恎傗桳塿側嵶嬠偑棙梡偱偒傞揝傪惂尷偡傞偙偲偵側傞丅偟偐偟丄崻寳偵偍偗傞揝偺棙梡壜擻惈傪惂尷偡傞偙偲偼丄堦斒揑偵丄崻寳偵惗懅偡傞嵶嬠偺昦尨惈傪懀恑偡傞婋尟惈偑偁傞丅傕偟偦傟偑丄戙幱偺嫤摨惈偲揝偺儗儀儖偑昦尨惈傪梷惂偡傞偙偲偑帵偝傟偰偄傞歁擕椶偺挵撪偺忬嫷偵帡偰偄傞偺偱偁傟偽丅塰梴僗僩儗僗墳摎偲怉暔柶塽宯偺枾愙側娭楢偼丄儕儞巁偵懳偡傞怉暔偺墳摎偱娤嶡偝傟偰偍傝丄塰梴妉摼偲怉暔柶塽墳摎偺堦斒揑偱暋嶨側棈傒崌偄傪帵嵈偟偰偄傞丅揝偲柶塽宯傪偮側偖僷僀僾栶偲偟偰丄IMA偼堦楢偺暋嶨偱懡柺揑側婡擻傪拠夘偡傞廳梫側惉暘偱偁傞傛偆偩丅堦曽偱偼丄IMA偺嬊強暘夝偵傛偭偰崻偺巁惈壔偑梷惂偝傟丄偦傟偵傛偭偰惉挿掆巭傗MAPK偺姰慡側儕儞巁壔側偳丄flg22偵懳偡傞崻偺墳摎偑壜擻偵側傞丅偙傟傜偺僨乕僞偲堦抳偡傞傛偆偵丄IMA偑寚懝偡傞偲丄崻偺昞柺偵忢嵼偡傞忢嵼嬠偺僐儘僯乕宍惉偑梷惂偝傟偨丅堦曽丄IMA1夁忚敪尰姅偼丄梩柺嶶晍偝傟偨P. syringae pv. tomato DC3000偵懳偟偰傛傝掞峈惈偱偁偭偨丅崱屻丄揝偺妉摼偲挋憼偺挷愡偑丄怉暔偺崻傗夎偵偍偗傞儅僀僋儘僶僀僆乕儉偺峔惉傗旝惗暔偺昦尨惈偵偳偺掱搙塭嬁偡傞偐傪挷傋傞偙偲偼嫽枴怺偄丅
丂丂
埲壓尨挊榑暥偺恾侾乣恾6傑偱偺慡暥傪栿偟偰偍偔偑丄
巻柺偺惂栺備偊偵丄恾侾丆恾2偺堦晹丄恾5偺堦晹偺傒傪奼戝揮嵹偟偰偍偄偨丅
丂
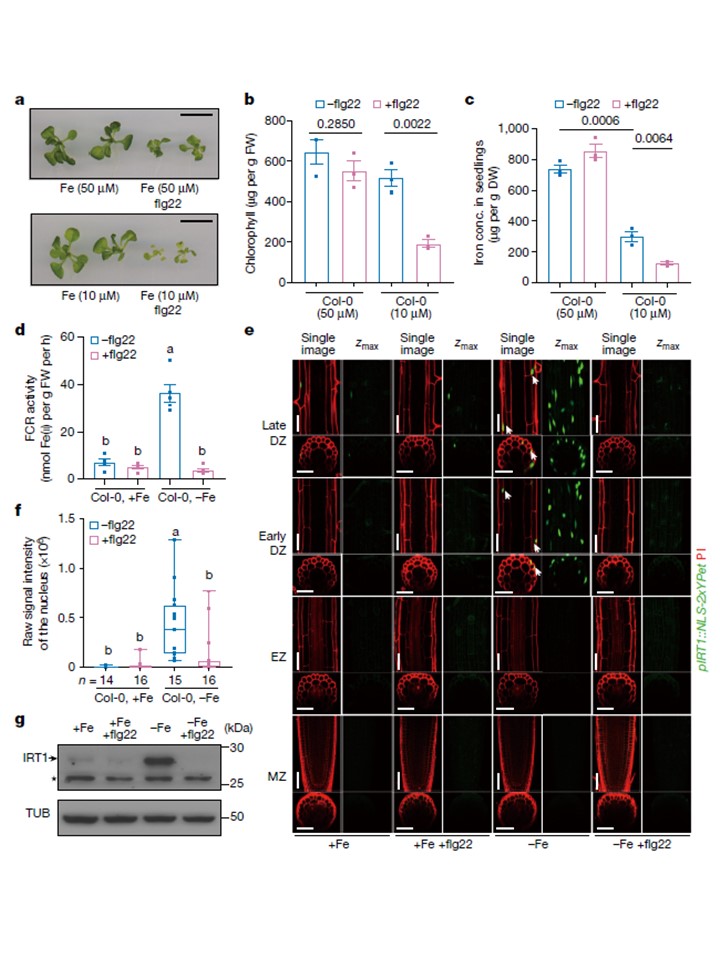
恾1乥flg22偼揝寚朢帪偵揝偺庢傝崬傒傪梷惂偡傞丅
a, 抧忋晹丅僗働乕儖僶乕丄cm丅
b, Col-0偺僔儏乕僩偺憤僋儘儘僼傿儖擹搙丅
c, Col-0昪偺揝擹搙丅3惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.丅摑寁夝愅偼椉懁t専掕傪梡偄偰峴偭偨丅DW, 姡憞廳検丅
d,+Fe忦審壓偱7擔娫惗堢偝偣偨Col-0崻偺FCR妶惈丅d, +Fe, +Fe + flg22, -Fe 傑偨偼 -Fe + flg22 塼懱攟抧偵 2 擔娫堏怉偟偨 Col-0 偺崻偺 FCR 妶惈丅5夞偺惗暔妛揑斀暅丅 僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅乮ANOVA乯傪梡偄偰暘愅偟偨堎側傞忦審娫偺摑寁揑偵桳堄側嵎傪帵偡丅
e,+Fe偵斀墳偟偨pIRT1::NLS-2xYpet昪偺崻偵偍偗傞IRT1僾儘儌乕僞乕妶惈丅昪偼+Fe 攟抧偱惗堢偝偣丄5擔屻偵暿偺塼懱攟抧偵堏偟丄24帪娫張棟偟偨丅DZ偼暘壔懷丄EZ偼怢挿懷丄MZ偼暘楐懷丅椢偼妀嬊嵼Ypet丄愒偼儓僂壔僾儘僺僕僂儉乮PI乯嵶朎暻愼怓丅奺張抲偵偮偄偰丄戙昞揑側扨堦嫟徟揰愗曅乮扨堦夋憸丄GFP 偍傛傃PI)丄嵟戝嫮搙z搳塭(zmax丄GFP偺傒)丄墶曽岦帇偺扨堦岝妛愗曅偍傛傃z 墶曽岦抐柺偺岝妛揑抐柺丄偍傛傃墶曽岦抐柺偺z-搳塭傪帵偡丅
f, pIRT1儗億乕僞乕偺惗偺僔僌僫儖嫮搙偺掕検壔丅堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅傪梡偄偰暘愅偟偨堎側傞忦審娫偺摑寁揑桳堄嵎傪帵偡丅堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偲Tukey偺専掕乮P < 0.05乯傪梡偄偰暘愅偟偨丅
g, +Fe, -Fe + flg22, -Fe 偍傛傃 -Fe + flg22 偺忦審壓偱惗堢偝偣偨 Col-0 偺崻偵偍偗傞 IRT1 僞儞僷僋幙儗儀儖偺僂僃僗僞儞僽儘僢僩暘愅丅栴報偼IRT僞儞僷僋幙僶儞僪傪帵偡丅 惎報偼旕摿堎揑側僶儞僪傪帵偡丅僠儏乕僽儕儞乮TUB乯僞儞僷僋幙偼撪晹僐儞僩儘乕儖偱偁傞丅
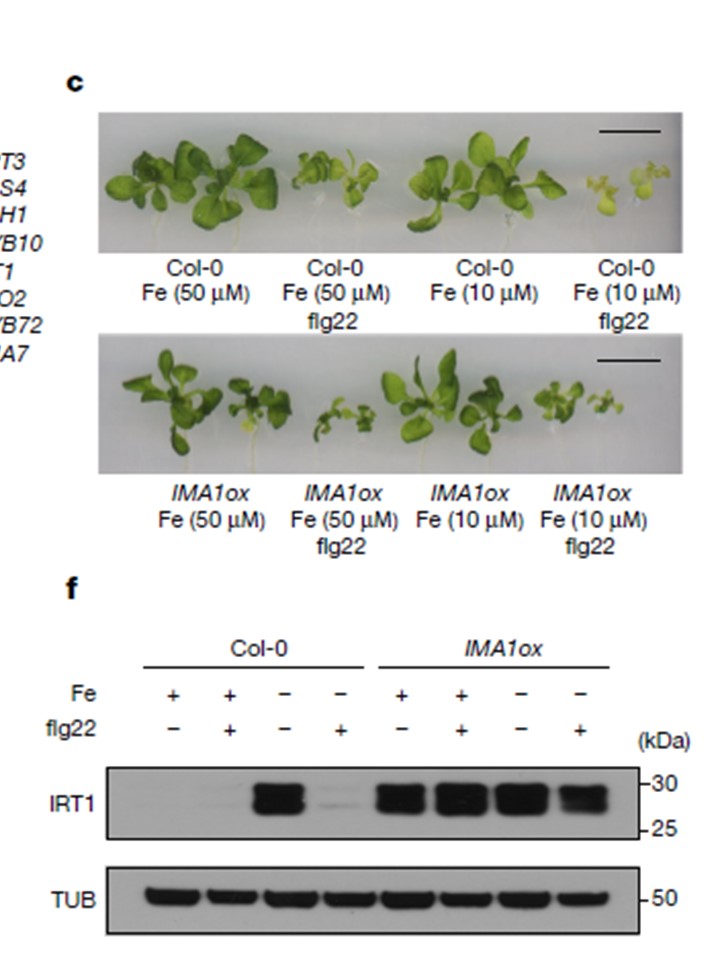
恾2乥flg22偼IMA1傪夘偟偰揝寚朢墳摎傪梷惂偡傞丅
a, DEGs偺RNA-seq夝愅丅
b, 傛偔抦傜傟偨揝墳摎惈堚揱巕乮3偮偺撈棫偟偨惗暔妛揑斀暅乯偺暯嬒拞怱z僗僐傾丗揝寚朢僔僌僫儖揱払惉暘丄揝庢傝崬傒惉暘丄挿嫍棧僔僌僫儖揱払惉暘丅
c-e,15擔楊偺Col-0偲IMA1ox偺昪偵偍偄偰丄掅儗儀儖偺flg22張棟乮10 nM乯偺桳柍偵偐偐傢傜偢丄揝暘廩懌乮50 兪M乯傑偨偼掅揝暘乮10 兪M乯忦審偵墳摎偟偨昞尰宆丅
c, 怴夎丅d, 夎偺憤僋儘儘僼傿儖擹搙 e, 昪偺揝擹搙丅d偲e偼3惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅P抣偼丄椉懁僗僠儏乕僨儞僩偺t専掕傪梡偄偰寁嶼偟偨丅
f, Col-0偍傛傃IMA1ox偺崻偵偍偗傞IRT1僞儞僷僋幙儗儀儖偺僂僃僗僞儞僽儘僢僩暘愅乮+Fe, +Fe + flg22, -Fe 偍傛傃 -Fe + flg22張棟偵墳摎偟偰乯丅
g, +Fe 忦審壓偱 7 擔娫惗堢偝偣丄+Fe, +Fe + flg22, -Fe 偍傛傃 -Fe + flg22 塼懱攟抧偵 2 擔娫堏怉偟偨 Col-0 偍傛傃 IMA1ox 偺崻偵偍偗傞 FCR 妶惈偺掕検暘愅丅5惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅堎側傞暥帤偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯屻偵Tukey偺専掕乮P < 0.05乯傪梡偄偰暘愅偟偨丄堎側傞忦審娫偺摑寁揑偵桳堄側嵎堎傪帵偡丅
恾3乥flg22偼崻偺抧忋慻怐偱IMA1傪嬻娫揑偵梷惂偡傞丅
a,b,pIMA1::eYFP-IMA1;ima8x偺崻偵偍偗傞IMA1僞儞僷僋幙儗儀儖丅撪晹僐儞僩儘乕儖偼僠儏乕僽儕儞丅
a, +Fe, +Fe + flg22, -Fe, -Fe + flg22張棟偵懳偡傞斀墳丅
b, 帪娫宱夁丗昪傪-Fe偱36帪娫慜張棟偟偨屻丄-Fe傑偨偼-Fe亄flg22乮1兪m flg22乯偱0丄3丄6帪娫張棟偟偨丅
c, 崻偺暘壔椞堟偵偍偗傞+Fe, +Fe + flg22, -Fe, -Fe + flg22張棟偵懳偡傞IMA1偺暘晍丅5擔楊偺IMA1::eYFP-IMA1;ima8x昪傪+Fe攟抧偱惗堢偝偣偨屻丄塼懱張棟攟抧偵24帪娫堏偟偨丅墿怓偼eYFP-IMA1丄愒怓偼PI嵶朎暻愼怓丅奺張棟偵偮偄偰丄戙昞揑側扨堦嫟徟揰愗曅乮扨堦夋憸丄eYFP/PI乯丄z僗僞僢僋偺嵟戝z搳塭乮zmax丄eYFP偺傒乯丄墶抐柺偺扨堦岝妛愗曅丄墶抐柺偺z搳塭傪帵偡丅僗働乕儖僶乕偼50兪m丅
d, pIMA1::eYFP-IMA1;ima8x偺崻偺暘壔椞堟偵偍偗傞IMA1寀岝僔僌僫儖嫮搙偺掕検壔丅摑寁夝愅偼丄椉懁Kruskal-Wallis専掕偵懕偒丄椉懁Steel-Dwass-Critchlow-Fligner庤弴傪梡偄偨懡廳堦懳斾妑傪梡偄偰峴偭偨乮P < 0.05乯丅
e,惓婯壔僔僌僫儖宎偺掕検朄偺柾幃恾丅
f, pIMA1::eYFP-IMA1;ima8x崻偺暘壔椞堟偵偍偗傞堎側傞張棟忦審壓偱偺惓婯壔IMA1僔僌僫儖宎偺掕検丅摑寁夝愅偼丄椉懁Kruskal-Wallis専掕偵懕偒丄椉懁Steel-Dwass-Critchlow-Fligner庤弴傪梡偄偨懡廳堦懳斾妑傪梡偄偰峴偭偨乮P < 0.05乯丅
g,h, IMA1敪尰傪嬱摦偡傞堎側傞慻怐摿堎揑僾儘儌乕僞乕傪帩偮僩儔儞僗僕僃僯僢僋怉暔偺崻偵偍偗傞IRT1僞儞僷僋幙儗儀儖偺僂僃僗僞儞僽儘僢僩暘愅丅撪晹僐儞僩儘乕儖偼僠儏乕僽儕儞偱偁偭偨丅
g, 掅揝偵傛傞IRT1偺撪場惈桿摫傪旔偗傞偨傔丄偡傋偰偺昪傪亄揝偱惗堢偝偣偨丅奺僩儔儞僗僕僃僯僢僋怉暔偺3偮偺撈棫偟偨宯摑傪帵偡乮Extended Data Fig.乯
h, pPGP4::mCtirine-IMA1 偺崻傪 +Fe 偍傛傃 偍傛傃 +Fe + flg22 偱偺張棟偵懳偡傞 pPGP4::mCtirine-IMA1 偺崻偺斀墳丅
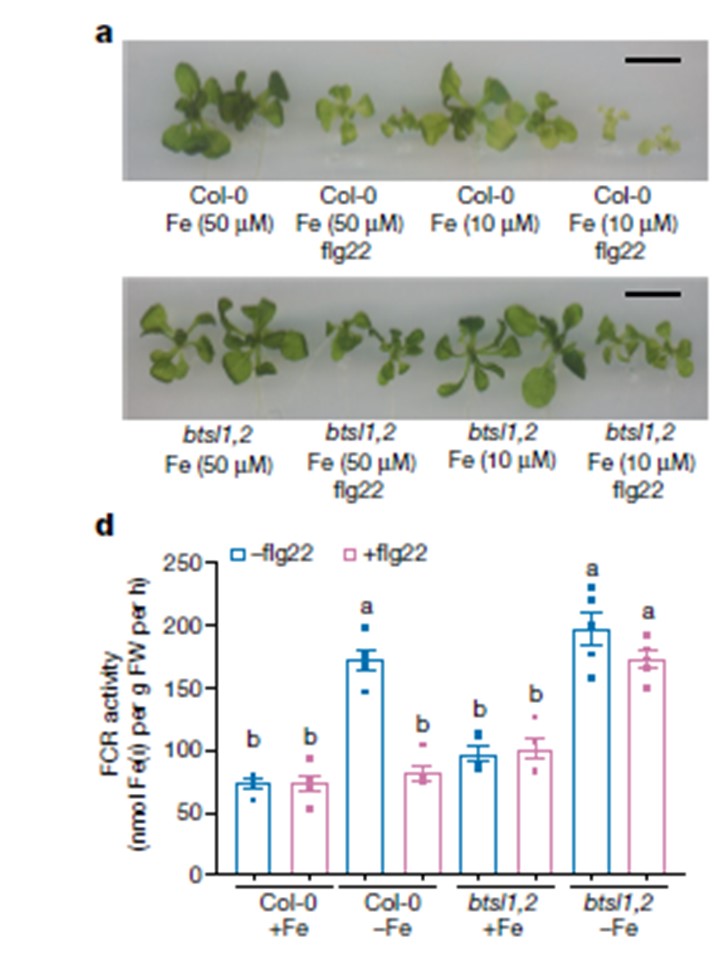
恾4乥flg22偼BTSL1偍傛傃BTSL2傪夘偟偨IMA1暘夝偵傛偭偰揝寚朢墳摎傪梷惂偡傞丅
a- c, +Fe乮50兪M乯傑偨偼-Fe乮10兪M乯拞丄flg22乮10nM乯柍張棟傑偨偼掅擹搙張棟偟偨15擔楊偺昪丅
b- a, 抧忋晹丅僗働乕儖僶乕偼1cm丅
b, 抧忋晹偺憤僋儘儘僼傿儖擹搙丅
c, 梒怉暔偺揝擹搙丅b偲c偼3惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅P抣偼椉懁僗僠儏乕僨儞僩偺t専掕傪梡偄偰寁嶼偟偨丅
d, +Fe 攟抧偱 7 擔娫惗堢偝偣丄+Fe, +Fe + flg22, -Fe 偍傛傃 -Fe + flg22 塼懱攟抧偵 2 擔娫堏怉偟偨 Col-0 偍傛傃 btsl1,2 偺崻偵偍偗傞 FCR 妶惈丅5惗暔妛揑暋惢丅
e, +Fe, +Fe + flg22, -Fe, -Fe + flg22張棟孮偺Col-0偍傛傃btsl1,2崻偵偍偗傞IRT1僞儞僷僋幙儗儀儖偺僂僃僗僞儞僽儘僢僩夝愅丅懳徠偼僠儏乕僽儕儞偱偁偭偨丅
f, +Fe, +Fe + flg22, -Fe, -Fe + flg22張棟偵懳偡傞pBTSL1-GFP偍傛傃pBSTL2-GFP偺崻暘壔懷偺嫟徟揰尠旝嬀墶抐柺丅椢丄GFP僠儍僱儖丅僗働乕儖僶乕丄50兪m丅
g, +Fe, +Fe + flg22, -Fe, -Fe + flg22張棟孮偺崻暘壔椞堟偵偍偗傞IMA1偺嫟徟揰尠旝嬀憸丅pIMA1::eYFP-IMA1;ima8x偍傛傃pIMA1::eYFP-IMA1;btsl1,2偺昪乮5擔楊乯傪+Fe攟抧偱惗堢偝偣偨屻丄塼懱張棟攟抧偵24帪娫堏怉偟偨丅墿怓偼eYFP-IMA1丄愒怓偼PI嵶朎暻愼怓丅張棟偛偲偵丄戙昞揑側嫟徟揰愗曅乮扨堦夋憸丄eYFP偲PI乯丄嵟戝z搳塭乮zmax丄eYFP偺傒乯丄墶抐柺恾偺扨堦岝妛愗曅丄墶抐柺恾偺z搳塭傪帵偡丅僗働乕儖僶乕偼50兪m丅
h,i, pIMA1::eYFP-IMA1;btsl1,2怉暔偺崻暘壔椞堟偵偍偗傞IMA1寀岝僔僌僫儖嫮搙(h)偲惓婯壔IMA1僔僌僫儖捈宎(i)傪丄+Fe丄+Fe + flg22丄-Fe偍傛傃-Fe + flg22張棟孮偱斾妑偟偨丅pIMA1::eYFP-IMA1;ima8x偼摨帪偵嶣塭偟偨偨傔丄恾3e,f偲摨偠僨乕僞僙僢僩傪巊梡偟偨丅摑寁 摑寁夝愅偼丄椉懁Kruskal-Wallis専掕偵懕偒 椉懁Steel-Dwass-Critchlow-Fligner朄傪梡偄偨乮P < 0.05乯丅
恾5乥IMA1偵埶懚偟偨崻寳巁惈壔偼丄崻偺柶塽墳摎偲嵶嬠僐儘僯乕宍惉傪惂屼偡傞丅
a,b, Col-0丄ima8x偍傛傃UBQ10::mCitrine-IMA1怉暔偺+Fe偍傛傃-Fe張棟屻偺崻寳巁惈壔斀墳偺夋憸壔乮a乯偍傛傃掕検壔乮b乯丅pH巜帵栻偲偟偰僽儘儌僋儗僝乕儖僷乕僾儖傪梡偄偨丅
b, +Fe丄+Fe+flg22丄-Fe偍傛傃-Fe+flg22張棟偵懳偡傞Col-0丄ima8x偍傛傃UBQ10::mCitrine-IMA1怉暔偵傛傞昗弨攟抧偺巁惈壔丅6惗暔妛揑斀暅丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅摑寁夝愅乮奺堚揱巕宆撪乯偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偵Tukey偺専掕傪壛偊偰峴偭偨乮P < 0.05乯丅椉懁Student偺t専掕偼丄+Fe忦審壓偱偺Col-0偲UBQ10::mCitrine-IMA1怉暔娫偺攟抧巁惈壔偺摑寁揑桳堄嵎傪帵偡偨傔偵梡偄偨丅
c,d丄MES-KOH柍揧壛乮c乯偍傛傃1mM MES-KOH揧壛乮d乯偺+Fe偍傛傃+Fe+flg22忦審壓偱偺Col-0丄ima8x偍傛傃UBQ10::mCitrine-IMA1怉暔偺崻偺挿偝偺掕検丅P抣偼丄椉懁僗僠儏乕僨儞僩偺t専掕傪梡偄偰寁嶼偟偨丅
e, Col-0丄ima8x偍傛傃UBQ10::mCitrine-IMA1怉暔懱偺MES-KOH偺桳柍偵傛傞flg22傪夘偟偨崻偺惉挿墳摎偺掕検乮+flg22/-flg22丄+flg22偺崻偺挿偝傪-flg22偺暯嬒偱妱偭偨傕偺乯丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅P抣偼椉懁僗僠儏乕僨儞僩偺t専掕傪梡偄偰寁嶼偟偨丅
f, +Fe 偍傛傃 +Fe +flg22 孮偵偍偗傞 CYP71A12 偺揮幨儗儀儖偼丄媡揮幨偵傛傞 qPCR乮RT-qPCR乯傪梡偄偰暘愅偟偨丅崻傪1兪M flg22偱1帪娫張棟偟偨丅敪尰抣偼ACT2偱惓婯壔偟偨丅3偮偺惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偍傛傃Tukey偺専掕乮P < 0.05乯傪梡偄偰峴偭偨丅
g, Col-0丄ima8x偍傛傃UBQ10::mCitrine-IMA1偺崻偵偍偗傞flg22偵傛傞MAPK儕儞巁壔偺僂僃僗僞儞僽儘僢僩夝愅丅崻偼1兪M flg22偱0丄5丄10暘娫張棟偟偨丅撪晹僐儞僩儘乕儖偼僠儏乕僽儕儞偱偁偭偨丅
h, 7擔楊偺僔儘僀僰僫僘僫偺崻傪丄+揝丄-揝丄偍傛傃旕棙梡壜擻揝乮nAvFe乯忦審壓偱丄CHA0偵傛傝愙庬1擔屻偵僐儘僯乕宍惉偝偣偨丅3惗暔妛揑暋惢丅僨乕僞偼暯嬒抣亇s.e.m.偱偁傞丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偵Tukey偺専掕傪壛偊偰峴偭偨乮P < 0.05乯丅
恾6乥IMA1偺拁愊偼丄抧昞偵惐懅偡傞嵶嬠傑偨偼怤擖偡傞嵶嬠偵傛偭偰柧椖偵挷愡偝傟傞丅
a, +Fe, +Fe + CHA0, -Fe, -Fe +CHA0, -Fe + flg22張棟孮偵偍偗傞pIRT1::NLS-2xYpet偺崻丅愒偼嵶朎昞柺偵嬊嵼偡傞CHA0-mCherry嬠丄椢偼妀偵嬊嵼偡傞Ypet丅戙昞揑側扨堦嫟徟揰愗曅乮扨堦夋憸丄GFP偲mCherry乯丄z僗僞僢僋偺嵟戝z搳塭乮zmax丄GFP偺傒乯丄偍傛傃墶抐柺偺扨堦岝妛愗曅傪帵偡丅僗働乕儖僶乕偼50兪m丅
b, +Fe, +Fe + CHA0, -Fe, -Fe + CHA0, -Fe + flg22張棟孮偺pIRT1::NLS-2xYpet崻偵偍偗傞惓婯壔僔僌僫儖掕検丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偍傛傃僼傿僢僔儍乕乮Fisher乯偺嵟彫桳堄嵎乮LSD乯専掕乮椉懁丄P < 0.05乯傪梡偄偰峴偭偨丅
c, pIMA1::eYFP-IMA1;ima8x怉暔偺崻暘壔椞堟偵偍偗傞+Fe, +Fe + CHA0, -Fe, -Fe + CHA0, -Fe + flg22張棟孮偵偍偗傞IMA1偺暘晍丅愒偼崻偺昞柺偵嬊嵼偡傞CHA0-mCherry嬠丄墿怓偼eYFP-IMA1乮墿怓僠儍儞僱儖乯丅戙昞揑側嫟徟揰愗曅乮扨堦夋憸丄eYFP偍傛傃mCherry乯丄z僗僞僢僋偺嵟戝z搳塭乮zmax丄eYFP偺傒乯丄偍傛傃墶抐柺偺扨堦岝妛愗曅傪帵偡丅僗働乕儖僶乕偼50兪m丅
d丄亄Fe丄亄Fe亄CHA0丄-Fe丄-Fe亄CHA0偍傛傃-Fe亄flg22張抲孮偵偍偗傞惓婯壔IMA1僔僌僫儖嫮搙丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偲僼傿僢僔儍乕偺LSD専掕乮椉懁丄P < 0.05乯傪梡偄偰峴偭偨丅
e, +Fe, +Fe + CHA0, -Fe, -Fe + CHA0, -Fe + flg22張棟孮偵偍偗傞昞旂-旂幙乮epi-cortex乯嵶朎憌偺惓婯壔IMA1僔僌僫儖嫮搙丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偲僼傿僢僔儍乕偺LSD専掕乮椉懁丄P < 0.05乯傪梡偄偰峴偭偨丅
f, +Fe, +Fe + CHA0, -Fe, -Fe + CHA0, -Fe + flg22搳梌孮偵偍偗傞惓婯壔IMA1僔僌僫儖宎偺掕検丅摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偲僼傿僢僔儍乕偺LSD専掕乮椉懁丄P < 0.05乯傪梡偄偰峴偭偨丅
g, +Fe, +Fe + CHA0, -Fe, -Fe + CHA0張棟孮偵偍偗傞丄懁崻尨婎偵偍偗傞eYFP-IMA1偺戙昞夋憸丅愒怓偼撪晹偵嬊嵼偡傞 CHA0-mCherry嬠丄墿怓偼eYFP-IMA1僔僌僫儖丅惎報偼 CHA0-mCherry偼懁崻巒尨晹埵偐傜崻偵怤擖偟偨丅栴報偼旂幙偵偍偗傞庛偄IMA1僔僌僫儖傪帵偡丅僗働乕儖僶乕偼50兪m丅扨堦 嫟徟揰愗曅乮扨堦夋憸丄eYFP偲mCherry乯丄z僗僞僢僋偺嵟戝z搳塭乮zmax z-僗僞僢僋偺嵟戝z-搳塭乮zmax丄eYFP偺傒乯丄偍傛傃墶曽岦帇偺扨堦岝妛愗曅傪帵偡丅BF丄柧帇栰丅僗働乕儖僶乕偼50兪m丅懁崻尨婎偵偍偗傞亄Fe丄亄Fe亄CHA0丄-Fe丄-Fe亄CHA0偍傛傃-Fe亄flg22張棟孮偵偍偗傞惓婯壔IMA1僔僌僫儖嫮搙偺掕検壔丅摑寁夝愅偼 摑寁夝愅偼丄堦尦攝抲暘嶶暘愅乮one-way ANOVA乯偲僼傿僢僔儍乕偺LSD専掕乮椉懁丄P < 0.05乯傪梡偄偰峴偭偨丅
恾1
恾2偺堦晹偺奼戝
恾4偺堦晹偺奼戝