総説:アラビドプシスのbHLH転写因子
アラビドプシスのbHLH転写因子
The arabidopsis bHLH transcription factor family
Fei Gao and Christian Dubos
Trends in Plant Science, Month 2023, Vol. xx, No. xx https://doi.org/10.1016/j.tplants.2023.11.022
要旨
塩基性らせん-ループ-へリックス(bHLHs)はすべての真核生物に存在し、植物に見られる転写因子(TFs)の最大のファミリーの一つを形成している。bHLHsは、環境との相互作用の中で植物の成長や発達に関わる重要なプロセス(例えば、気孔や根毛の発達、鉄の恒常性、暑さや日陰への応答など)に関与する遺伝子の転写活性化因子および/または抑制因子として機能している。最近の研究では、複雑な制御ネットワークにおけるbHLH TFsの機能についての理解が深まりつつあり、そこでは一連の翻訳後修飾(PTM)が、細胞内局在、DNA結合能、転写活性、安定性(タンパク質間相互作用、リン酸化、ユビキチン化、スモイル化など)を制御する上で重要な役割を担っている。bHLHの機能と制御をさらに解明することは、一般的な植物の生物学の理解を深め、作物改良のための新しいツールを開発するのに役立つであろう。
図2. シロイヌナズナ(Arabidopsis thaliana)における塩基性ヘリックス-ループ-ヘリックス(bHLH)転写因子(TF)が関与するシグナル伝達経路の概略図。(A)気孔の発達には、2つの異なるサブファミリー(すなわちIaとIIIe)に属する5つのbHLHの連続的な活性が必要である。(B)根毛の発生は、3つの異なるサブファミリー(すなわちIIIf、VIIIc、XI)に属する12個のbHLHによって制御されている。非毛細胞(atrichoblast)では、MBW複合体がGL2(GLABARA2、ホメオドメインTF)の発現を活性化し、その結果、根毛の発達に関与するbHLHの発現が抑制される。有毛細胞(三葉芽細胞)では、MBW複合体内でCAPRICE(CPC/R3-MYB)がWERWOLF(WER/R2R3-MYB)に取って代わり、根毛の発達に関与するbHLHの発現を可能にする。(C)高温に応答して、PHYTOCHROME-INTERACTING FACTOR 4(PIF4;サブファミリーVII)が誘導され、FLOWERING TIME(FT)と成長関連遺伝子の発現を活性化し、それぞれ開花と細胞伸長を促進する。しかし、PIF4はSPEECHLESS(SPCH/bHLH98;サブファミリーIa)の発現を抑制し、それによって気孔の生産を阻害する。(D)日陰に応答した細胞伸長は、4つの異なるサブファミリー(すなわち、Va、VI、VII、XII)に属する15個のbHLHの活性によって促進され、その活性は(b)HLH(VII、XVIIa、XVIIbサブファミリー由来)との相互作用によって阻害される。これらの相互作用はもう一つの(b)HLHs(サブファミリーXV)によって拮抗されることがあり、これらの(b)HLHsは日陰誘導性の細胞伸長を可能にする二次層抑制因子として働く。略号 ACE, ACTIVATOR FOR CELL ELONGATION; BEE, BR ENHANCED EXPRESSION; BIM, BES1-INTERACTING MYC-LIKE; PRE, PACLOBUTRAZOL RESISTANCE; TTG1, TRANSPARENT TESTA GLABRA 1 (WD40-repeat protein).
図3. シロイヌナズナ(Arabidopsis thalian)の鉄ホメオスタシスを制御する塩基性ヘリックス-ループ-ヘリックス(bHLH)制御ネットワークの簡略図。18種類のbHLH転写因子 Ib、IIIa、IIIe、IIIf、IVa、IVb、IVcの7つのサブファミリーからなる18種類のbHLH転写因子が、複雑な制御ネットワークの中で働き、鉄の恒常性を制御している。略語とサブファミリー:bHLH Ib:bHLH38, bHLH39, bHLH100, および bHLH101; bHLH IVa:bHLH18, bHLH19、
bHLH20、およびbHLH25;bHLH IVc: IAA-LEUCINE RESISTANT 3(ILR3/bHLH105)、IRON DEFICIENCY TOLERANT 1(IDT1/bHLH34 bHLH34)、bHLH104、およびbHLH115;FIT、FER-LIKE IRON DEFICIENCY INDUCED TRANSCRIPTION FACTOR;MYC1/(bHLH12)。bHLH12: サブファミリーIIIf; MYC2/bHLH6: サブファミリーIIIe; PYE, POPEYE.
図4. シロイヌナズナ(Arabidopsis thaliana)における塩基性ヘリックス-ループ-ヘリックス(bHLH)転写因子(TF)を制御する翻訳後メカニズム。二量体化は、bHLH転写因子の活性を制御する主な翻訳後修飾の一つであり、(A)ホモ二量体(例えば、鉄ホメオスタシスの制御;図3参照)または(B)ヘテロ二量体(例えば、分裂組織における細胞増殖の制御)のいずれかである。(C)拮抗的なbHLH/(b)HLH相互作用は、bHLHヘテロ二量化のもう一つの例である(例えば、陰影に応答した細胞伸長の制御)。(D)bHLH以外のタンパク質もbHLHと直接相互作用して、その活性を調節することができる(例えば、ジベレリンシグナルDELLAリプレッサーによる鉄ホメオスタシスの調節)。(E)bHLH以外のタイプのTFとの相互作用も、その活性に影響を与える(例えば、MBWタンパク質複合体の形成による、種皮におけるプロアントシアニジン生合成の制御;図2B参照)。翻訳後マークの付加もまたbHLHの活性と安定性を調節する。
その中で、(F)リン酸化[例えば、CBL-INTERACTING PROTEIN KINASE 11 (CIPK11)プロテインキナーゼによるFER-LIKE IRON DEFICIENCY INDUCED TRANSCRIPTION FACTOR (FIT)のリン酸化は鉄の取り込みを促進する]、 (G)ユビキチン化[例えば、bHLH115とIAA-LEUCINE RESISTANT 3(ILR3)のユビキチン化依存性分解による鉄の取り込み阻害]、(H)スモイル化[例えば、MYC2のスモイル化によるPLANT U-BOX 10(PUB10)との相互作用の阻害、ひいてはその分解と光形態形成制御における役割の促進]が最も研究されている。bHLHの略語とサブファミリー: ACE, ACTIVATOR FOR CELL ELONGATION (subfamily XII); BTS, BRUTUS LIKE (E3-ubiquitin ligase); GL3/bHLH1, GLABROUS 3 (subfamily IIIf); EGL3/bHLH2, ENHANCER OF GLABROUS 3 (subfamily IIIf); LHW/bHLH156, LHL1/bHLH173, そして LHL2/bHLH155 (bHLH subfamily XIII); IBH1/bHLH158(bHLHサブファミリーXVIIa);PRE1/bHLH136、PACLOBUTRAZOL RESISTANCE(bHLHサブファミリーXV);PUB10、PLANT U-BOX10(E3-ユビキチンリガーゼ);TMO5/bHLH32、T5L1/bHLH30、およびT5L2/bHLH107: (bHLHサブファミリーVb);TT2(R2R3-MYB);TT8/bHLH42、TRANSPARENT TESTA 8(サブファミリーIIIf);TTG1、TRANSPARENT TESTA GLABRA 1(WD-リピートタンパク質)。
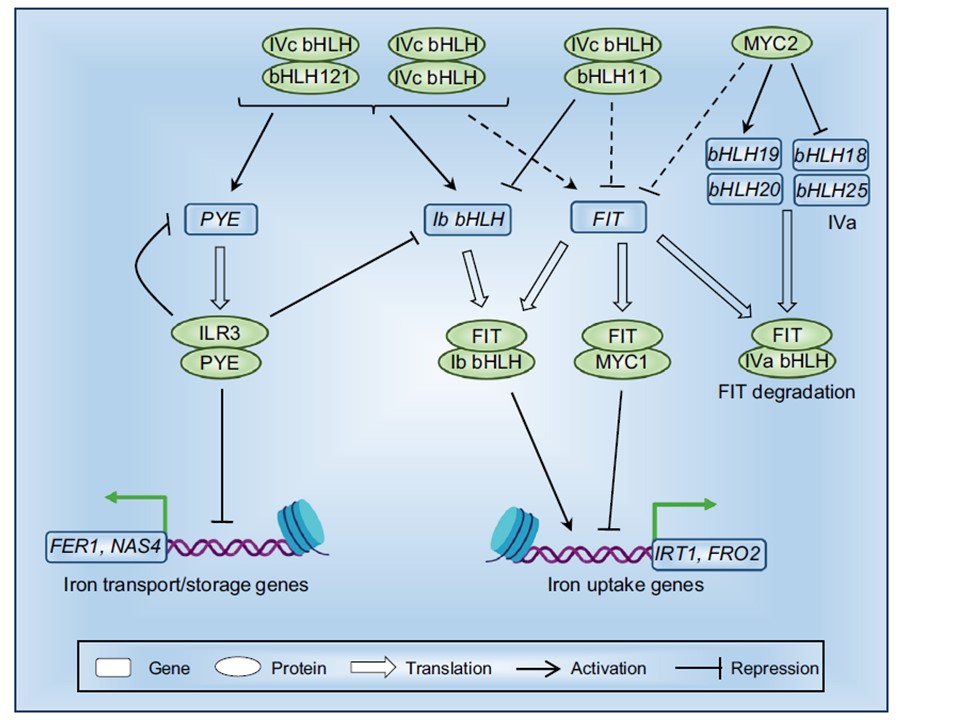
図2
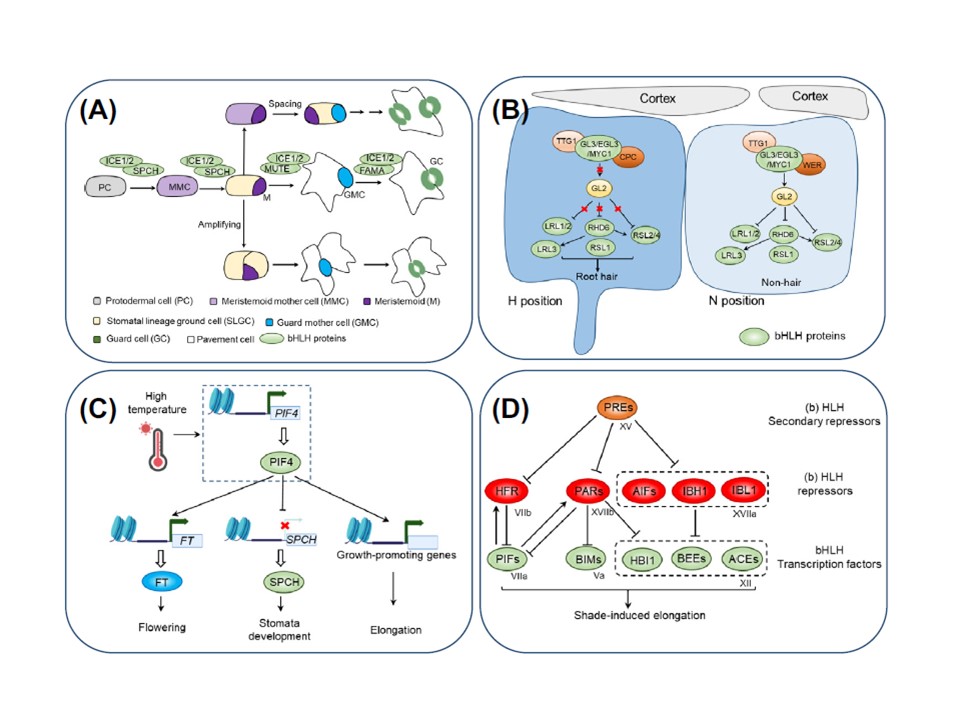
図3
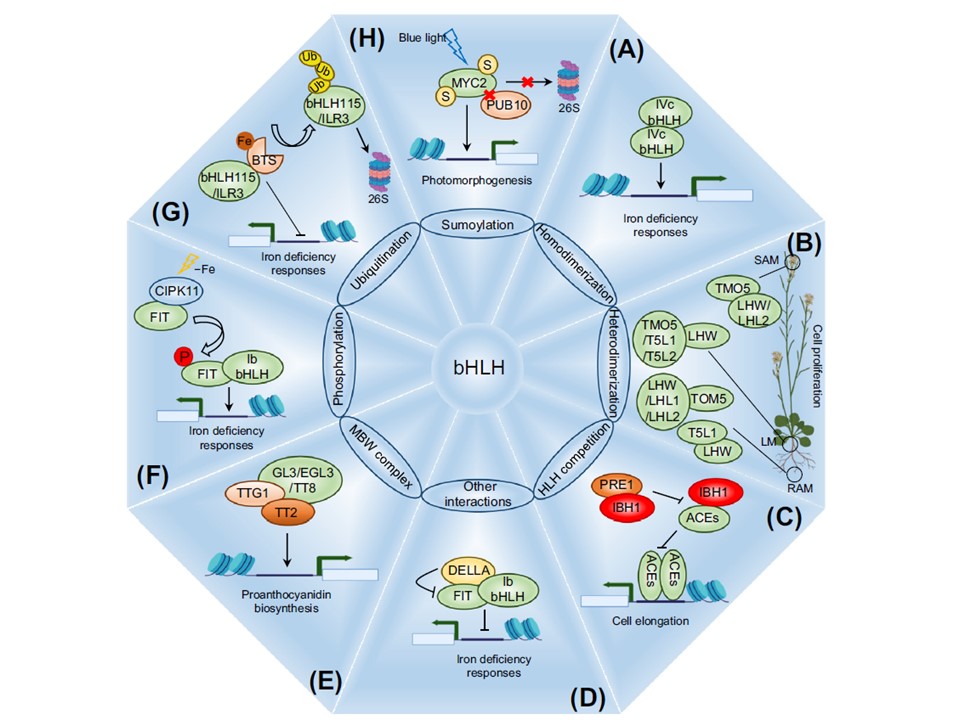
図4