植物はどのように競合する利害を調整するのか
このGuerinotによる解説文は、鞭毛タンパクfragerrinのペプチド断片が植物体内のIMA,BTSを介して、植物の根からの鉄吸収を抑止する、というきれいなストーリーを描いている。これが真実なら、病原菌により感染すると植物が鉄欠乏になることを説明するものである。ただし感染菌が鞭毛を有している場合に限る。次回にはCaoらのnatureの原著論文を紹介するつもりである。
植物はどのように競合する利害を調整するのか
How plants iron out competing interests
Shanice S. Webster & Mary Lou Guerinot
https://doi.org/10.1038/d41586-023-03995-3
News & views
(要旨)植物が病原体を認識すると、その防御戦略の一環として鉄の摂取を控える。そのメカニズムには、鉄の取り込み経路を活性化する分子を分解することで、根が鉄を獲得するのを抑制することが含まれる。
(以下、全訳した)
植物はどのようにして、鉄の取り込みを調節して自らの成長を促進し、関連する無害な微生物(常在微生物)に栄養を与える一方で、病原微生物を抑制するというバランスをとっているのだろうか?Nature誌に掲載されたこの論文(Cao, M. et al. Nature https://doi.org/10.1038/s41586-023-06891-y (2024))は、鉄欠乏時における鉄と免疫のクロストークの分子メカニズムに関する洞察を提供し、植物とその常在細菌および病原性細菌との間の複雑な相互作用を浮き彫りにしている。
鉄は、植物とそれに付随する微生物にとって不可欠な微量栄養素である。電子伝達を促進するその能力は 光合成や呼吸のような主要な細胞プロセスにとって非常に貴重なものであるが、過剰量の鉄は有害である。そのため、植物は鉄の取り込みをコントロールし、適切なレベルを維持しなければならない。この鉄のホメオスタシスにより、植物は悪影響を最小限に抑えながら、恩恵を最大限に享受することができる。さらに、鉄は病気の原因となる微生物の増殖を助ける可能性があるため、植物も動物と同様に、防御戦略として鉄の利用を制限するメカニズムを進化させてきた。
鉄のホメオスタシスは植物の免疫防御と関連しているが、鉄欠乏と植物の免疫防御シグナルのクロストークのメカニズムについてはほとんど知られていない。
Caoらは、鞭毛と呼ばれる細菌表面の構成要素であるフラジェリンタンパク質由来のペプチド断片であるflg22が、ペプチドIMA1が分解される過程を通して鉄の取り込みを抑制することを示した。IMA1は、移動性シグナルペプチドであるIMAファミリーの1つである。これらは顕花植物全体で進化的に保存されており、鉄欠乏時に発現し、鉄欠乏反応を活性化するのに必要な転写因子タンパク質のターンオーバーを妨げることによって、鉄の取り込みを制御する。著者らは、鉄が欠乏した植物では、鉄の取り込みに重要な2つのタンパク質-FRO2と呼ばれる酵素と、鉄輸送体タンパク質-RT1が、flg22の存在下では誘導されないことを示している(図1)。
Caoらは、遺伝子発現をモニターするトランスクリプトーム解析という手法を用いて、これらの発見を確認した。その結果、鉄欠乏によって強く誘導され、flg22への細胞暴露によって抑制される遺伝子群が明らかになった。注目すべきは、それらの遺伝子の中にIMAファミリー(IMA1、IMA2、IMA3)のメンバーが含まれていたことである。そこで著者らは、IMAはおそらくflg22シグナル伝達カスケードの下流で機能しているのではないかと考えた。著者らは、IMA1のレベルが通常よりも高い植物変異体を用いて、flg22による鉄の取り込み抑制が消失することを見いだした。これは、IMA1の関与をさらに裏付けるものであり、flg22によるIMA1の機能への直接的な干渉を示唆するものである。flg22がIMA1の機能をどのように制御しているかを調べるために、著者らはIMA1がどこで発現しているかを調べた。Caoたちは、IMA1が根の分化領域と呼ばれる領域に局在し、表皮と皮層と呼ばれる細胞タイプに存在することを発見した。IMA1の発現は、flg22処理によってこれらの場所で減少した。著者らは、FRO2とIRT1の発現を誘導するためには、IMA1が皮層と表皮で局所的に発現している必要があると提唱している。Caoたちは、flg22がIMA1を枯渇させるメカニズムを明らかにしようとした。IMAペプチドは、BTSタンパク質によってユビキチンというタンパク質が付加されることで修飾されることが知られており、修飾されたIMAペプチドはその後、プロテアソームと呼ばれる分解装置によって分解される。BTSタンパク質は主に地上部と、植物の根の内側の内梢と呼ばれる構造で発現している。しかし、BTSファミリーの他の2つのメンバー、BTSL1とBTSL2は根で発現し、鉄レベルを調節する。Caoらは、野生型植物と比較して、遺伝子btsl1およびbtsl2(BTSL1およびBTSL2をコードする)に変異を持つ植物は、鉄欠乏の徴候が少なく、flg22免疫活性化防御応答に対する感受性が低いことを示している。さらに、btsl1/btsl2変異株では、鉄欠乏下でIMA1タンパク質のレベルが上昇したことから、flg22によるIMA1の抑制はBTSL1/2依存的に起こることが示唆された。flg22がどのようにしてIMA1の分解につながるのか?この点については未解明のままであるため、BTSL1/2依存的なIMA1の破壊を媒介するものを見つける探求が続けられている。
植物の根は、微生物にとって魅力的なニッチとなる。IMA1が免疫と鉄の取り込みに二重の役割を担っていることを考えると、このタンパク質の制御は微生物のコロニー形成に顕著な影響を与える可能性がある。興味深いことに、Caoらは、IMA1が高発現している植物では、常在菌であるシュードモナス・プロテゲンス(旧名シュードモナス・フルオレッセンス)によるコロニー形成が増加する一方、病原性細菌であるシュードモナス・シリンゲによるコロニー形成は減少することを示している。しかし、両菌種に鞭毛が存在することから、なぜ常在菌と病原菌の鞭毛に対して植物が異なる反応を示すのかが疑問視されている。いくつかの研究室がこの疑問に取り組んでいる。常在細菌は多くの多様なflg22配列をコードしており、そのほとんどは免疫系に認識されない。また、根が局所的な抗菌免疫反応を起こすには、微生物特異的タンパク質と植物組織へのダメージの両方が必要であり、常在菌を温存しながら病原菌に対応する効果的な戦略が明らかになったようだ。今後は、マイクロバイオームコミュニティの文脈におけるIMA1の役割を探る必要があるだろう。
Cao氏らの研究結果は、植物が病原体を検出すると、鉄分が十分な条件下で鉄分の取り込みを制限するのと同じ成分のいくつかを使って、鉄分を控えるプログラムを実行できることを説明するのに役立つ。BTSタンパク質は鉄摂取経路のゲートキーパーであるため、病原菌がBTSを標的にするのは当然かもしれない。P. syringaeの一部の株が産生する細菌タンパク質AvrRps4は、BTSを封鎖し、病原体が利用できる鉄の取り込みを可能にする。
鉄の場合と同様に、リン酸飢餓応答と植物免疫の間にもクロストークがある。これらのメカニズムは、栄養素の獲得と免疫反応との間に、基本的かつおそらく一般的なつながりがあることを浮き彫りにしている。我々は、微生物と宿主の間の栄養素をめぐる競争に関する理解を利用して、植物が病原菌に対する感受性を不用意に高めることなく、食品の栄養素含有量を高める持続可能な解決策を開発する必要がある。植物が病原菌の繁殖を抑えながら、どのようにして栄養分の取り込みのバランスをとっているのか、科学者たちが今後さらに解明していくことになるだろう。
図1 の説明
a,植物が鉄を必要とするとき、土壌からの鉄の取り込みはIMA1というタンパク質によって仲介され、鉄取り込み複合体(青)を形成するIRT1とFRO2というタンパク質の生産を促進する。IMA1は根の外側の細胞層(表皮と皮層)に存在するが、内皮と呼ばれる内側の層には存在しない。FLS2レセプターに結合する細菌ペプチドflg22がない場合、タンパク質BTSL1とBTSL2はIMA1を標的としない。
b,鉄は有害なバクテリアの成長を助ける可能性があり、Caoらは、バクテリアが感染している間、植物がどのように鉄の取り込みを制限しているかに光を当てた。flg22がFLS2に結合して活性化すると、BTSL1とBTSL2の作用によってIMA1が分解され、鉄取り込み複合体が失われる。
シャニース・S・ウェブスターは生物学部、ハワード・ヒューズ医学博士である。ハワード・ヒューズ医学研究所 7710, USA。
Mary Lou Guerinotは 生物科学科に在籍、ダートマス大学、ハノーバー、ニューハンプシャー州 03755、USA。
電子メール:shanice.webster@duke.edu;
guerinot@dartmouth.edu
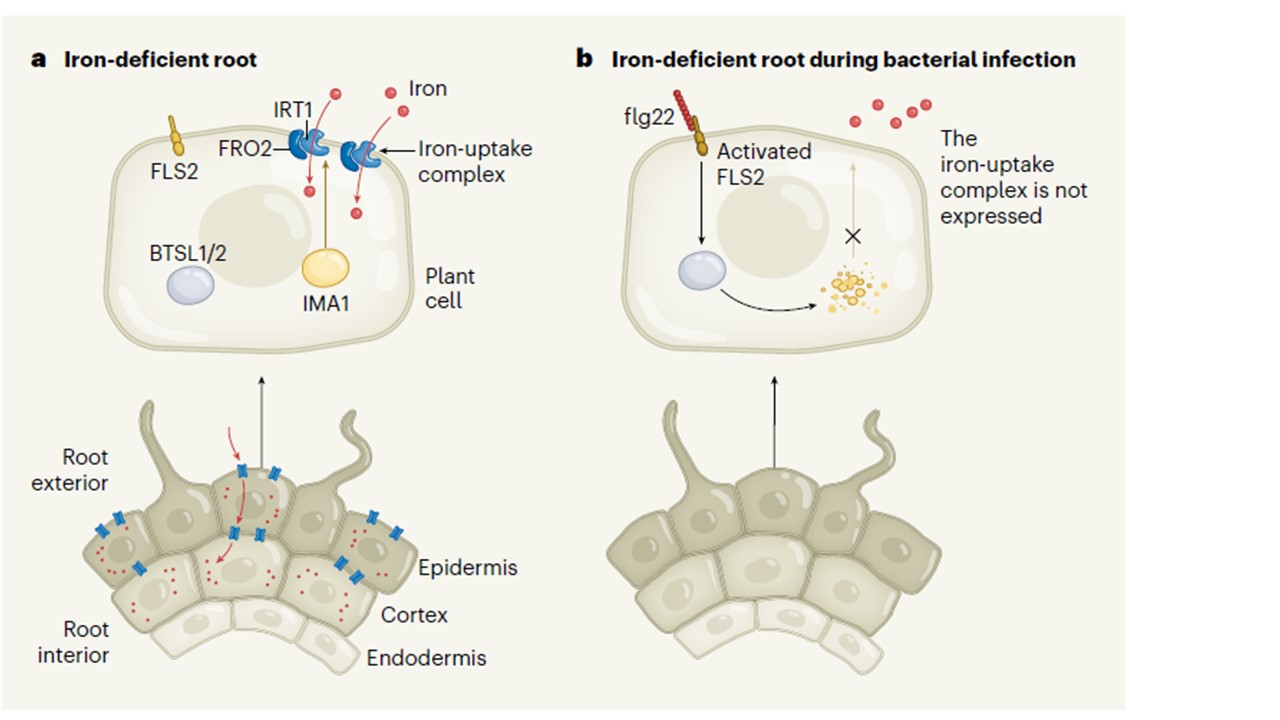
図1