揝偺儂儊僆僗僞僔僗偲僇僪儈僂儉掞峈惈偵偍偗傞IMA儁僾僠僪偺婡擻
偙傟傑偱IMA儁僾僠僪偲僇僪儈僂儉偺娭學偵偮偄偰扤傕尵媦偟偰偙側偐偭偨偲巚偆丅偙偺榑暥偼偦偺寠応傪慱偭偨傕偺偲尵偊傛偆丅
揝偺儂儊僆僗僞僔僗偲僇僪儈僂儉掞峈惈偵偍偗傞IMA儁僾僠僪偺婡擻
IMA peptides function in iron homeostasis and cadmium resistance
Ruonan Wang a,b , Yuchen Fei a,b , Yilin Pan a,b , Peijun Zhou a,b , Julius Oluwaseun Adegoke a,b , Renfang Shen a,b , Ping Lan a,b,* a State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China b University of Chinese Academy of Sciences, Beijing 100049, China
丂丂丂
Plant Science 336 (2023) 111868
丂丂
乮梫栺乯
昁恵旝検塰梴慺偱偁傞揝乮Fe乯偼丄岝崌惉丄屇媧丄偦偺懠懡偔偺峺慺斀墳偵娭梌偟偰偄傞丅
懳徠揑偵丄僇僪儈僂儉乮Cd乯偼帠幚忋偡傋偰偺惗暔偵偲偭偰桳撆側尦慺偱偁傞丅揝偺寚朢偲
僇僪儈僂儉偺撆惈偼丄擾嶌暔偺惉挿偲惗嶻惈傪挊偟偔懝側偄丄偮偄偵偼恖娫偺寬峃栤戣偵傑偱敪揥偡傞丅
怉暔偑偳偺傛偆偵揝偺庢傝崬傒偲儂儊僆僗僞僔僗傪惂屼偟丄僇僪儈僂儉撆惈偲摤偭偰偄傞偺偐傪棟夝偡傞偙偲偼丄揝傪朙晉偵娷傓偑僇僪儈僂儉偼彍嫀偝傟偨泱庫傪恖娫偺偨傔偵奐敪偡傞偨傔偵昁恵偱偁傞丅僔儘僀僰僫僘僫傗僀僱傪梡偄偨嵟嬤偺尋媶偱丄埲壓偺偙偲偑柧傜偐偵側偭偨丅
IRON MAN (IMA)儁僾僠僪偑丄儐價僉僠儞偲嫞崌揑偵憡屳嶌梡偡傞偙偲偵傛偭偰丄揝寚朢偵懳墳偡傞偨傔偺廳梫側挷愡場巕偱偁傞偙偲傪柧傜偐偵偟偨丅
儐價僉僠儞E3儕僈乕僛偲嫞崌揑偵憡屳嶌梡偡傞偙偲偱丄IVc僒僽僌儖乕僾偺bHLH揮幨場巕乮TFs乯偺暘夝傪慾奞偡傞丅
IMA偺崅敪尰偼丄僔儘僀僰僫僘僫偍傛傃僐儉僊偺椉曽偱丄揝寚朢傪妶惈壔偡傞偙偲偵傛傝僇僪儈僂儉僗僩儗僗偵懳偡傞懴惈傪梌偊傞 丅偙偙偱偼丄IMA儁僾僠僪偑揝寚朢墳摎偵偍偄偰婡擻偟丄揝偺儂儊僆僗僞僔僗傪払惉偟丄僇僪儈僂儉撆惈偲摤偆僼傽僀僩儗儊僨傿僄乕僔儑儞偺愽嵼揑側岓曗偱偁傞偲偄偆偙偲傪媍榑偡傞丅
乮埲壓偼恾偺愢柧乯
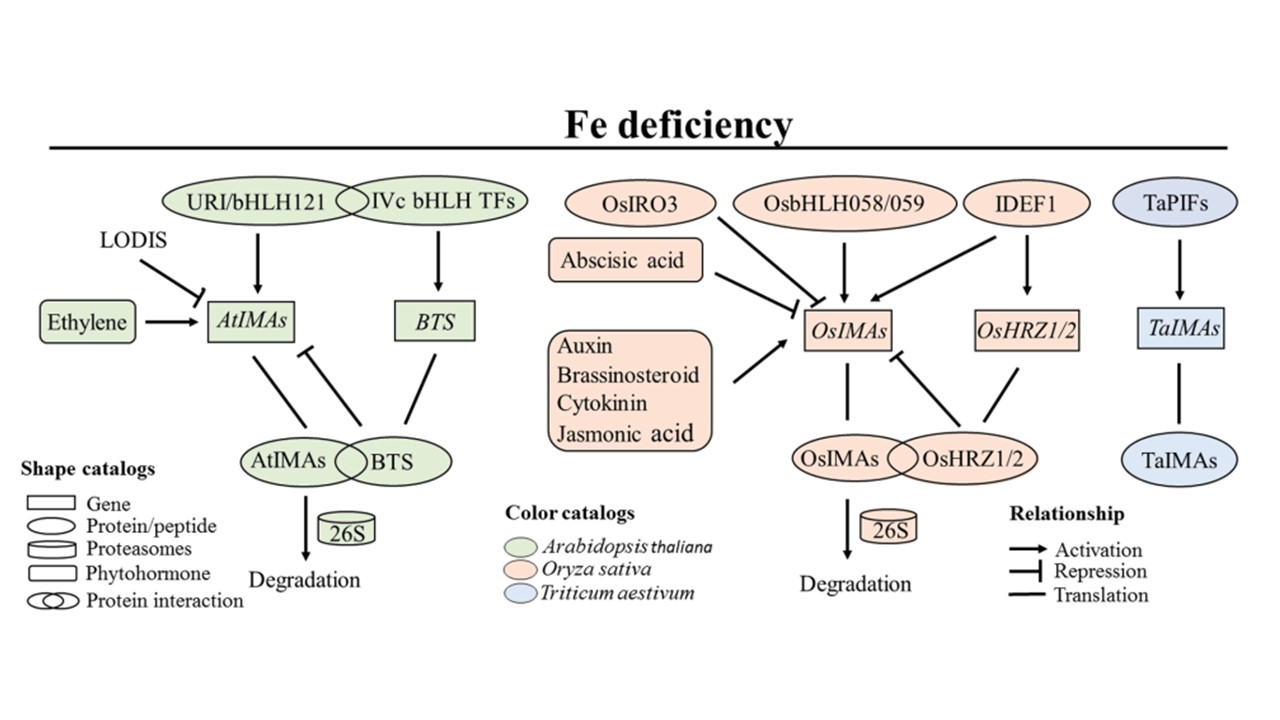
恾2. IMA儁僾僠僪堚揱巕妶惈偺惂屼丅
揝暘偺廩懌壓偱偼丄IMA儁僾僠僪堚揱巕偺婎慴敪尰偼旕忢偵掅偄偐専弌尷奅埲壓偱偁傞偨傔丄恾偵偼帵偟偰偄側偄丅IMA儁僾僠僪堚揱巕偺敪尰偼丄僔儘僀僰僫僘僫乮嵍乯偲僀僱乮拞墰乯偱偼寑揑偵桿摫偝傟丄僐儉僊乮塃乯偱偼傢偢偐偩偑桳堄偵桿摫偝傟傞丅僔儘僀僰僫僘僫偱偼丄AtIMA偺敪尰偼庡偵URI/bHLH121偲IVc bHLH TF偺1偮偲偺憡屳嶌梡偵傛偭偰惂屼偝傟偰偄傞丅僄僠儗儞偼億僕僥傿僽偵丄LODIS乮挿嫍棧揝僔僌僫儖乯偲BTS偼僱僈僥傿僽偵AtIMA偺敪尰傪揮幨惂屼偡傞丅AtIMA儁僾僠僪偺僞乕儞僆乕僶乕偼丄26S僾儘僥傾僜乕儉儅僔儞傪夘偟偨E3儕僈乕僛BTS偵傛偭偰惂屼偝傟偰偄傞丅僀僱偱偼丄OsIMA偺敪尰偼庡偵OsbHLH058/059偵傛偭偰惂屼偝傟丄IDEF1偵傛偭偰惓偵惂屼偝傟傞偑丄OsIRO3偵傛偭偰晧偵惂屼偝傟傞丅僆乕僉僔儞丄僽儔僔僲僗僥儘僀僪丄僒僀僩僇僀僯儞丄僕儍僗儌儞巁偼OsIMA偺敪尰傪惓偵惂屼偡傞偑丄傾僽僔僕儞巁偲OsHRZ1/2偼OsIMA偺敪尰傪晧偵惂屼偡傞丅OsIMA儁僾僠僪偺僞乕儞僆乕僶乕偼丄26S僾儘僥傾僜乕儉傪夘偟偨E3儕僈乕僛OsHRZ1/2偵傛偭偰惂屼偝傟偰偄傞丅僐儉僊偱偼丄TaIMA偺敪尰偼TaPIF乮Phytochrome-Interacting Factor乯條僞儞僷僋幙偵傛偭偰揮幨儗儀儖偱惂屼偝傟偰偄傞偙偲偑曬崘偝傟偰偄傞丅
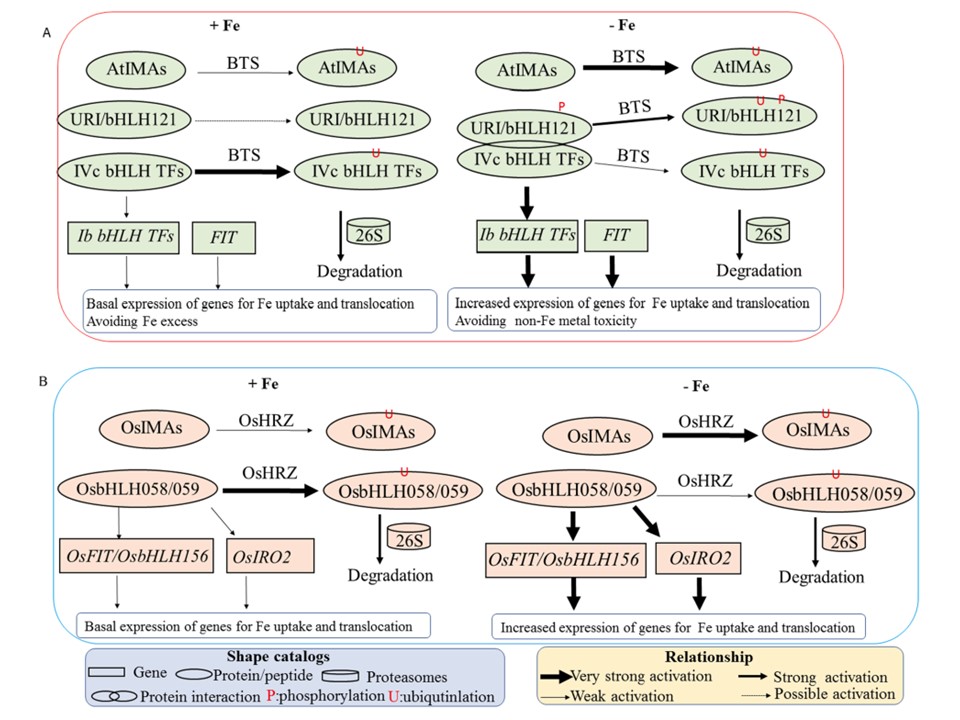
恾3.
MA儁僾僠僪偼揝寚朢壓偱揝儂儊僆僗僞僔僗挷愡僱僢僩儚乕僋偵娭梌偡傞丅Strategy-I乮A乯偲Atrategy-II乮B乯偺椉曽偵偍偄偰丄IMA儁僾僠僪偑揝挷愡僱僢僩儚乕僋偱摥偔扨弮側儌僨儖丅
娙扨偵尵偆偲丄IMA儁僾僠僪偼揝寚朢帪偵崻偵尠挊偵拁愊偟偨丅堦曽丄IVc bHLH TFs偺慡懱揑側敪尰偼丄僔儘僀僰僫僘僫丄僀僱偲傕偵揝偺忬懺偵偐偐傢傜偢曄壔偟側偐偭偨丅IMA儁僾僠僪傕IVc bHLH TFs傕丄傑偨僔儘僀僰僫僘僫偱偼儕儞巁壔偝傟偨URI傕丄僔儘僀僰僫僘僫偱偼BTS丄僀僱偱偼HRZ偲偄偆儐價僉僠儞E3儕僈乕僛偵傛偭偰僞乕儞僆乕僶乕偝傟丄嵟廔揑偵偼26S僾儘僥傾僜乕儉傪夘偟偰暘夝偝傟傞丅偙偺僞儞僷僋幙偺僞乕儞僆乕僶乕偼丄IMA儁僾僠僪偲E3儕僈乕僛偺嫞崌揑憡屳嶌梡偵傛偭偰朩偘傜傟丄IVc bHLH TF偼斾妑揑崅偄儗儀儖偵曐偨傟傞丅偦偺寢壥丄IVc bHLH TF乮僀僱偺僆儖僜儘僌OsbHLH58偲OsbHLH59乯偼丄僔儘僀僰僫僘僫偱偼儕儞巁壔偝傟偨URI/bHLH121乮僀僱偱偼儕儞巁壔偝傟偰偄側偄乯偲憡屳嶌梡偟丄壓棳偺Ib bHLH TF乮僀僱偺僆儖僜儘僌OsIRO2乯傗FIT乮僀僱偺僆儖僜儘僌OsbHLH156乯偺敪尰傪桿敪偟丄嵟廔揑偵揝偺庢傝崬傒偲堏摦偺堚揱巕偺敪尰傪桿摫偡傞丅
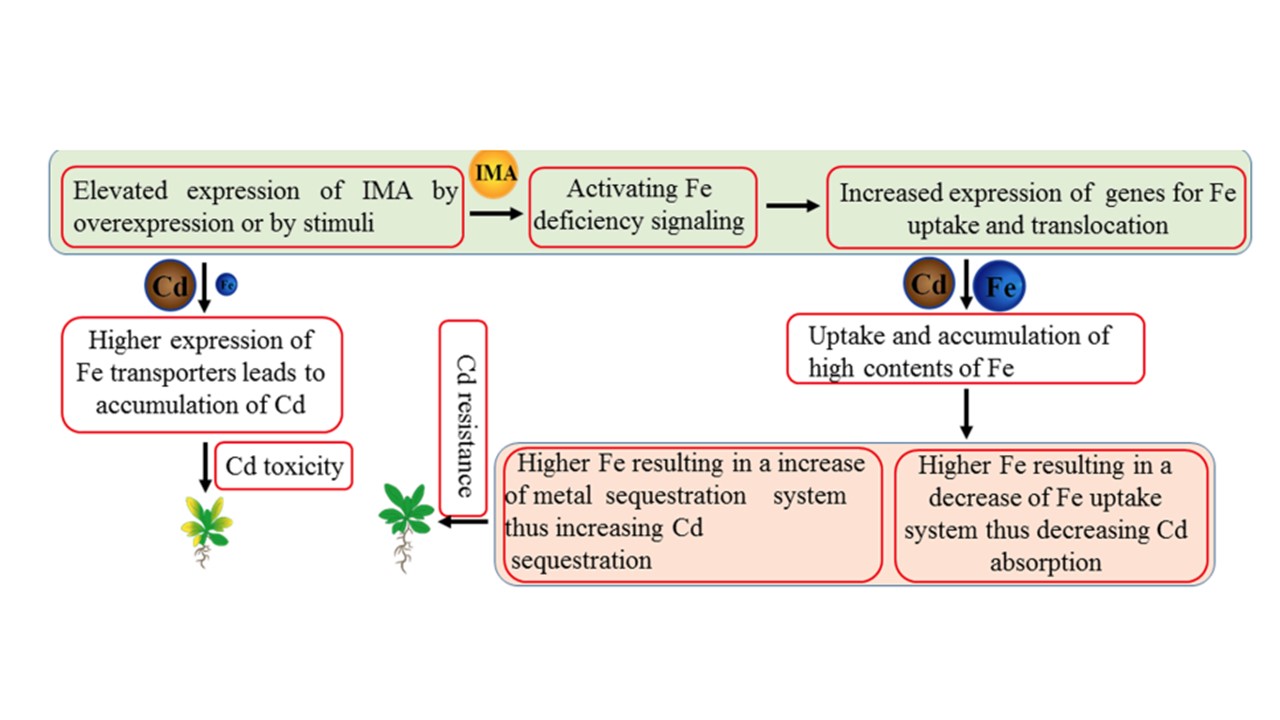
恾4.
IMA儁僾僠僪偺敪尰忋徃偑怉暔偵僇僪儈僂儉掞峈惈傪梌偊傞儌僨儖丅
IMA儁僾僠僪偺崅敪尰偼丄僔儘僀僰僫僘僫傗僀僱偱偼嫮椡側僾儘儌乕僞乕傪夘偟偨堎強惈敪尰偵傛偭偰丄僐儉僊偱偼扽慺揰(carbon dots)側偳偺壔妛揑巋寖偵傛偭偰丄Fe寚朢僔僌僫儖揱払偑妶惈壔偝傟丄偦偺寢壥丄Fe偺庢傝崬傒偲堏摦偵娭偡傞堚揱巕偺敪尰偑憹壛偡傞丅Fe庢傝崬傒堚揱巕偺敪尰偑崅傑傞偲丄惗堢攟抧偵Cd偑懚嵼偡傞偑Fe偑晄懌偡傞応崌偵丄Cd娷検偑拁愊偝傟丄Cd撆惈偑桿敪偝傟丄嵟廔揑偵怉暔偺惗堢偵僟儊乕僕傪梌偊傞偙偲偵側傞丅懳徠揑偵丄椉愴棯怉暔偵偍偗傞Fe庢傝崬傒堚揱巕偺崅敪尰偲丄愴棯II怉暔偵偍偗傞僼傿僩僔僨儘僼僅傾偺崅敪尰偼丄僇僪儈僂儉僗僩儗僗偑懚嵼偟偰傕丄惗堢攟抧偑Fe偱廫暘側応崌丄Fe娷検傪憹壛偝偣傞丅崅偄Fe娷検偼丄傗偑偰Fe庢傝崬傒僔僗僥儉傪掆巭偝偣丄僇僪儈僂儉偺媧廂傪尭彮偝偣傞丅堦曽丄崅偄揝暘偼揝妘棧僔僗僥儉傪妶惈壔偟丄僇僪儈僂儉偺妘棧傪憹壛偝偣傞丅憤偠偰丄僇僪儈僂儉媧廂偺尭彮偲僇僪儈僂儉妘棧偺憹壛偺慻傒崌傢偣偑丄僇僪儈僂儉掞峈惈偵婑梌偡傞丅
恾2
恾3
恾4