鉄の導管への負荷と長距離輸送に関する動的コンパートメントモデルは、シロイヌナズナにおけるカナマイシンの金属吸収への影響を説明する
WBC19はABCGファミリーの1/2サイズのATP結合カセットトランスポーターをコードしており、そのほとんどのメンバーが排出機能することが知られている。著者らはアラビドプシスのwbc19変異株にカナマイシン(Kan)を投与すると、極端に地上部への鉄や亜鉛などの重金属含量が低下することから、WBC19が導管への鉄・亜鉛などの負荷のための冗長トランスポーターではないかという仮説を検証しようとしている。この論文はそのモデル実験である。図表結果の図表が多いので省略して、(まえがき)と(考察)を全訳した。まえがきはよく書ていると思う。
鉄の導管への負荷と長距離輸送に関する動的コンパートメントモデルは、シロイヌナズナにおけるカナマイシンの金属吸収への影響を説明する
A dynamic compartment model for xylem loading and long-distance transport of iron explains the effect of kanamycin on metal uptake in Arabidopsis
Ayalew Mentewab、Bethany W. Mwaura、Carla M. Kumbale、Catherine Rono、Natalia Torres-Patarroyo、Toma´sˇ Vlcˇko、Ludmila Ohnoutkova´、Eberhard O. Voit
Front. Plant Sci. 14:1147598. doi: 10.3389/fpls.2023.1147598
(要旨)
抗生物質カナマイシン(Kan)に暴露されたシロイヌナズナは、金属ホメオスタシスが変化していることがわかった。
さらに、WBC19遺伝子の変異により、カナマイシンに対する感受性が高まり、鉄(Fe)および亜鉛(Zn)の取り込みが変化することがわかった。
ここでは、金属の取り込みとKanへの曝露の間のこの驚くべき関係を説明するモデルを提案する。
まず、金属摂取現象に関する知識を利用して、輸送と相互作用の図を作成し、それを基にダイナミックコンパートメントモデルを構築する。
このモデルでは、導管に鉄とそのキレーターを投入するための3つの経路を設定している。
1つは、未知のトランスポーターが関与する経路で、Feがクエン酸(Ci)とキレートを形成して導管に取り込まれる。
この輸送ステップは、Kanによって著しく阻害されることがある。
これと並行して、FRD3はCiを導管に輸送し、そこで遊離鉄とキレートを形成することができる。
第三の重要な経路は、WBC19が関与することである。WBC19は、金属-ニコチアナミン(NA)主にFe-NAキレートを、そしておそらくNAそのものも輸送する。
定量的な探索と分析を可能にするため、実験的な時系列データを用いて、この説明と予測モデルをパラメータ化した。
その数値解析により、二重変異体による反応を予測し、野生型、変異体、Kan阻害実験によるデータ間の観察された差異を説明することができた。
重要なことは、このモデルによって、植物が突然変異やカナマイシンによる鉄輸送阻害の影響に対抗するメカニズム的戦略をリバースエンジニアリングすることができ、金属ホメオスタシスに関する新しい知見が得られたことである。
(はじめに)
植物はミネラルを抽出するために土壌を採掘するが、その際にカナマイシン(Kan)のような抗生物質を取り込み、植物の正常な機能に悪影響を及ぼすことがある。Kanは、土壌細菌Streptomyces kanamyceticusが産生するアミノグリコシド系抗生物質である。アミノグリコシドは主に、原核生物および真核生物のリボソームの小サブユニットに結合することによって作用し、二次的にタンパク質合成を阻害する。
シロイヌナズナを含むほとんどの植物はKanに対して感受性が高く、この抗生物質はネオマイシンホスホトランスフェラーゼ(NPTII)マーカーを含むトランスジェニック植物の選抜に日常的に用いられている。このよく知られた感受性とは対照的に、WBC19(ABCG19)がKan耐性を与えることが発見された(Mentewab and Stewart, 2005)。
WBC19はABCGファミリーの1/2サイズのATP結合カセットトランスポーターをコードしており、そのほとんどのメンバーが排出機能することが知られている。ノックアウトしたシロイヌナズナのwbc19変異体はKanに対して強い感受性を示すが、WBC19を過剰発現させたトランスジェニック・タバコ植物は抵抗性を獲得した(Mentewab and Stewart, 2005)。現在までのところ、抵抗性の基礎となるメカニズムは不明であるが、複雑なようである。例えば、大腸菌でWBC19を発現させても、Kan抵抗性の有意な増加は見られなかった。さらに、wbc19プロモーター-GUS融合体を発現させたトランスジェニックシロイヌナズナでは、子葉で高レベルの発現が検出され、根、古い葉、花の維管束柔組織でも検出された。
これらの発現パターンから、WBC19が養分の長距離輸送に重要な役割を果たしていることが示唆される。Kanによる影響とWBC19の寄与を理解するため、シロイヌナズナのトランスクリプトームをKan曝露時に解析した。その結果、Kanが金属ホメオスタシスに影響を及ぼすことが示され、金属分析の結果、Kanが金属取り込みに有意な影響を及ぼすことが確認された。具体的には、対照植物がKanに曝露された場合、Feの取り込みは60%減少し、Cuレベルは50%増加した。この結果は、植物がKanに暴露された反応、または結果として、Feの取り込みが制限されることを示唆している。さらに、Kan非存在下で生育させた対照植物とwbc19変異体植物の金属レベルを比較したところ、ZnとCuレベルに有意な差が認められた。具体的には、変異株は対照株よりもZnとCuが約50%少なかったが、FeとMnのレベルは影響を受けなかった。これらの結果は、Kan非存在下でもWBC19が金属取り込みに関与していることを示唆している。
最後に、wbc19変異体では、鉄濃度は対照植物よりもさらに低く、CuとZnの濃度は最低に達し、Mnの濃度は著しく低くなった。このように、wbc19変異体は対照植物と同様に鉄の取り込みを減少させる反応を示し、全体的な金属含量の低下はKanへの暴露によって強く悪化した。これらの結果から、シロイヌナズナにおける抗生物質と金属ホメオスタシスの関連性が明らかになった。鉄と亜鉛は植物に最も多く含まれる微量栄養素のひとつであり、その取り込みと移動は高度に制御されている。シロイヌナズナの根の表皮細胞に鉄が入る主な経路は、FRO2/IRT1システムに基づいており、鉄キレート還元酵素2(FRO2)が鉄(III)を可溶性の鉄(II)に還元し、鉄調節輸送体1(IRT1)がその後に細胞内への鉄の通過を可能にする。根の表皮細胞に入った鉄は、プラスモデスマータを経由して維管束系へと移動し、Iron Regulated1/Ferroportin 1 (IREG1/FPN1)によって導管に運ばれると考えられている。
しかし、ireg1変異体では、鉄の輸送とシグナル伝達には大きな影響がないことから、冗長なトランスポーターの存在が示唆される。導管に入ると、鉄はクエン酸によってキレートされ、鉄(III)クエン酸(FeCi)複合体を形成し、長距離輸送を促進する(Larbi et al., 2010; Rellán-Alvarez et al., 2010)。このプロセスにとって重要なのは、根の形成体から導管へのCiの流出であり、これは多剤・毒素排出ファミリー(MATE)トランスポーターであるferric reductase deficient 3(FRD3)によって可能になる。FRD3は根の内鞘に発現し(Rogers and Guerinot, 2002; Green and Rogers, 2004)、木部柔細胞から導管にCiを輸送し、それによって植物全体にFeの長距離輸送を可能にする(Durrett et al., 2007)。ZnもIRT1や他のZIPファミリートランスポーターによって取り込まれる (Grotz et al., 1998; Vert et al., 2001; Connolly et al., 2002)。
一旦根のシンプラストに入ると、導管に取り込まれるか、根の表皮細胞から押し出される。導管への負荷は主に2つのP型ATPアーゼ、すなわち重金属ATPアーゼ2と4(HMA2とHMA4)によって達成される(Hussain et al.) 植物カドミウム抵抗性2(PCR2)は、Zn過剰の条件下で、導管におけるZnの負荷と根の表皮細胞からの排出の両方を仲介する(Song et al.)。 Znはまた、陽イオン拡散促進因子ファミリーの2つのトランスポーター、金属耐性タンパク質1および3(MTP1およびMTP3)を介して根の液胞に隔離されることができる(Kobaeら、2004;Desbrosses-Fonrougeら、2005;Arrivaultら、2006)。亜鉛キレーター、特にニコチアナ ミン(NA)の存在、輸送、コンパートメント化も、亜鉛の取り込みと 分布に重要な役割を果たしている(Sinclair and Krämer, 2012; Klatte et al.) CiもNAも、Fe、Zn、その他の金属をキレートする可能性を持っており、錯体の安定性はpHと化学量論に依存する(von Wirenら、1999)。したがって、導管の弱酸性のpHでは、Ciが鉄の主要なキレーターであり、細胞質の中性pHでは、NAが鉄の主要なキレーターであると考えられている。NAはまた、導管におけるZnとCuの主要なキレーターであると考えられている。金属ホメオスタシスの多くの構成要素、特に鉄と亜鉛の取り込みについては、その特徴が明らかにされている。
しかし、潜在的に冗長な鉄トランスポーターの正体や、導管にNAがどのように負荷されるかについては、まだギャップが残っている。さらに、WBC19の役割とKanの影響も、金属取り込みの理解にまだ組み込まれていない。
この目的のために、我々は、導管における鉄の取り込みが、まだ同定されていないFe-Ciトランスポーターと、Fe-NAや、おそらくZn-NAやNAそのものを含む他の金属-NA複合体を輸送すると思われるWBC19に依存しているという概念モデルを提案する。重要なのは、このモデルでは、Fe-CiトランスポーターはKanの存在下で阻害され、その時点でWBC19が主要なFeトランスポーターとして機能すると規定していることである。この提案の定量的解析を可能にするため、概念モデルを動的コンパートメントモデルに変換した。まず、シロイヌナズナにおける金属取り込みに関する一般的および特異的な知識を用いて、動的モデルの構築の基礎となる相互作用図を考案した。このモデルは、べき乗関数の形で阻害を許容する、わずかに一般化された質量作用系として定式化される(Voit, 2013; Voit et al.) この形式は間違いなく最も単純で偏りが少ない。
第2段階として、本論文の実験セクションで述べた時系列データを用いて、このモデルをパラメータ化する。これにより、野生型と突然変異および抑制実験から得られたデータの間に観察された差異を、植物がこれらの変化に反応する生理学的戦略という観点から解釈し、説明することができる。
4 考察
図1の図から明らかなように、様々な金属、キレーター、トランスポーターの作用や相互作用は、直感的にはほとんど理解できない。我々の知る限り、このアプローチにより、シロイヌナズナにおける金属負荷と輸送に関する初めての包括的モデルが生まれた。提案されたコンパートメントモデルは、野生型のシステムをよく捉えている。重要なことは、このモデルによって、植物が悪条件に効果的に対応するための代償戦略を探ることができたことである。過去に多くの著者が、特定の系における調節シグナルの推論を提案し、生物系における調節シグナルの不在または存在を最適化する系統的な方法さえ開発してきた(例えば、Hatzimanikatisら、1996a;Hatzimanikatisら、1996b)。
というのも、単なる有無ではなく、変化の大きさに興味があったし、植物は静止しているわけではなく、特に実験中に成長するからである。
ここで推測し、文書化した戦略は非独特的でアドホックなものであったが、この種のモデルを用いて、不利な状況を補う潜在的に効果的な戦略に関する新しい仮説を検証する方法を実証することができた。このモデルは、長距離鉄輸送に関するこれまでの理解に、2つの要素を加えるものである。それは、未知のFe-Ciトランスポーターと、Fe-NAトランスポーターとしてのWBC19(そしておそらく他の金属-NAまたはNAトランスポーター)である。Ciトランスポーターとして以前に確立されたFRD3とともに、このモデルは、3つのトランスポーターによって異なるコンパートメント間を移動する際の基質間の相互作用を捉えている。
このモデルでは、2つのトランスポーターがFeの輸送を可能にし、2つのトランスポーターがCiの輸送を可能にするという、ある程度の自然な冗長性を示している。しかし、各トランスポーターは、Fe-Ci、Fe-NA、Ciという異なる形態のFeとキレーターの移動を可能にするが、いずれも完全に等価ではない。さらに、これらのトランスポーターは、Feに加えて、他の金属キレートも輸送する可能性がある。我々のモデルは、様々な金属-キレーター複合体の化学種を特定しようとはしていない。なぜなら、これらは動的であり、どの特定の形態が輸送されるかについては、不確実性が大きいからである。全体として、これらのトランスポーターそれぞれを介した金属移動は、その存在、活性、基質に対する親和性、およびそれぞれの基質の存在量によって決定される。コンセプトの証明として、そしておそらくより高度な解析の出発点として、我々の計算モデルは鉄と亜鉛のみを考察している。明らかに、このモデルは、ここで提案した設計とパラメータ化に類似したプロセスで、他の金属に拡張することができる。ここではFeとZnに焦点をあてたが、これは植物に最も多く含まれる2つの微量栄養素であり、キレーターCiとNAとの親和性が実験によって研究されているからである(von Wiren et al.)
しかし、一見冗長な系を通る鉄輸送の詳細や、植物の制御反応は明らかではない。導管負荷の構成要素に加えて、我々のモデルは、局所的および全身的なシグナルによって媒介される鉄欠乏に対する既知の制御応答を統合している(Vert et al.) 提案されたモデルは、質的にも量的にも我々の実験結果と一致しており、標準的な条件下およびKanにさらされたときのwbc19およびfrd3変異体の表現型を再現している。特に、wbc19変異体の特徴的なKan感受性表現型や、frd3変異体における導管への鉄の蓄積増加と構成的な鉄欠乏反応を捉えている。このモデルの重要な点は、Fe-CiトランスポーターがKanによって阻害されるという事実である。このメカニズムは、wbc19変異体に対するKanの強い効果と、Kanに暴露された植物における鉄取り込みの全体的な減少を説明する(Mentewab et al.)。 さらに、frd3変異体に対するKanの強い効果も予測される。この結果は、Kanがfrd3変異体の表現型を悪化させるという我々の実験結果によって裏付けられた。KanによるFe-Ciトランスポーターの阻害の潜在的なメカニズムと意義は不明である。しかし、1つの可能性として、シロイヌナズナ植物がFe-Ciの輸送を制限するのは、同じトランスポーターを介したKanの輸送を防ぐためであると考えられる。
その結果、MAR1が変異すると、苗は抗生物質に対する抵抗性が増加する(Conte et al.) これと似たようなシナリオが、われわれの推定するFe-Ciトランスポーターにもあるのかもしれない。もしそうであれば、今回のモデルは、植物の栄養状態に過剰な影響を与えることなく、植物がKanの移動を制限することを可能にする微妙なメカニズムを表していることになる。ダイナミックモデルを使った新しい試みとして、我々はこのモデルを用いて、シロイヌナズナが変異体やKanによる阻害にどのように応答するかを説明した。補償がなければ、例えばwcb19突然変異はそれ自体でバイオマス中の鉄と亜鉛の量を大幅に減少させるだろう(表3)。しかし、我々が観察しているのはそうではない。突然変異やKanの阻害による機能不全を、どのように定量的に補うことができるかを直感的に判断するのは難しいだろう。このモデルによって、そのような補償のための潜在的なメカニズムを探ることができ、これらの代替補償戦略のいくつかを植物でテストすることは興味深い。結論として、我々はシロイヌナズナにおいて観察されたKanと金属取り込みの関係を説明するモデルを確立した。このモデルは、カナマイシンと様々な突然変異によって影響を受ける金属輸送に関する仮説を検証することができる。今後の研究では、未知の鉄-銅トランスポーターの同定に焦点を当てる予定である。
このモデルでは、Fe-Ciトランスポーターの変異は、WBC19がFe輸送の代替ルートになるため、Fe取り込みへの影響は限定的であると予測している。しかし、FRD3との二重変異体では、導管への遊離またはキレート化Ciの輸送が阻害されるため、そのような苗が生存可能であれば、frd3変異体よりも悪い表現型になるはずである。同様に、WBC19を持つ二重変異体では、導管への鉄輸送は事実上停止し、苗が生育可能であれば、強い鉄欠乏をもたらすであろう。対照的に、IREG1との二重変異体は強い表現型を示さないはずである。このモデルは、さらに他の金属を含むように拡張することができる。CiとNAがさまざまな金属をキレートすることを考えると、このモデルの構成要素は、CuやCoなどの金属の恒常性と交差するだろう。
図1
シロイヌナズナにおける鉄および亜鉛の取り込み系の経路系図と変数名。
WBC19がZn-NAまたはNAを輸送する能力はモデルにとって重要ではないため、灰色の線で示した。
同様に、当初は考えられていたが、最終的には重要でない制御シグナルも灰色で示した。
図2
50mg/lのKanを添加または無添加のMS培地で生育させた対照およびwbc19苗の総鉄および亜鉛含量の時間経過。点とエラーバーは、3反復から平均した実験測定値とその標準偏差を示す。線はモデル結果を示す。
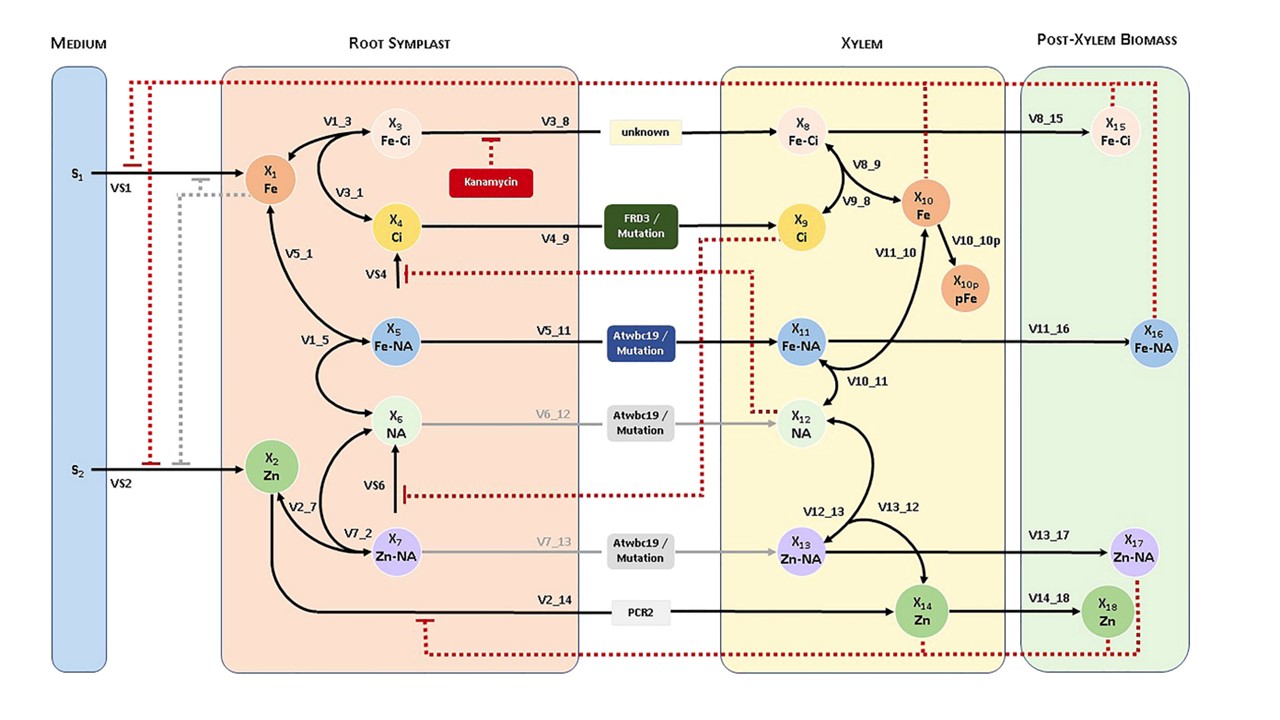
図1

表1
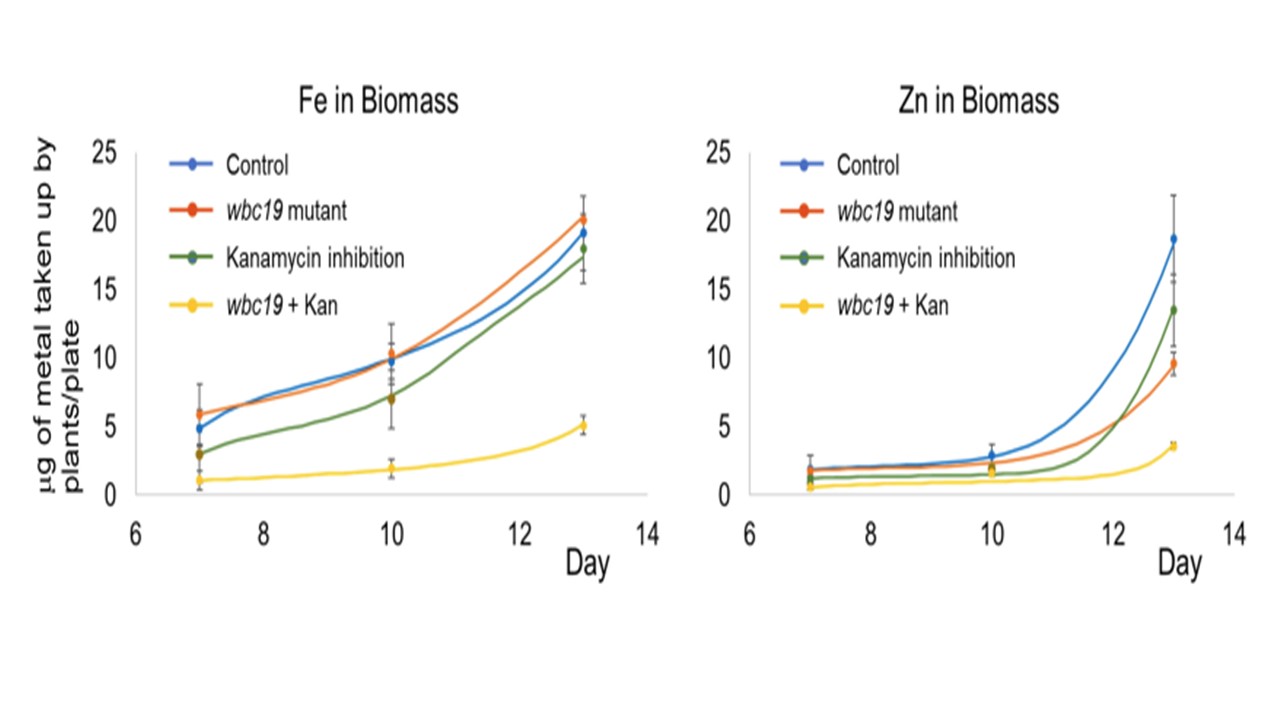
図2