ヒストン変異体HTB4はIb bHLH転写因子を介した鉄ホメオスタシスのエピジェネティック制御により葉の老化を遅らせる
以下は中国本土の4つの研究機関の共同研究である。エピジェネテイック変異ー>転写因子ー>鉄ホメオスタシス制御ー>葉の老化 という、一連の遺伝子制御の流れを解明したきわめてオリジナルなな研究である。
葉の老化制御における[HTB4]-[bHLH TFs]-[FRO2/IRT1]-[Feホメオスタシス]というシグナル伝達カスケードを提案している。
ヒストン変異体HTB4はIb bHLH転写因子を介した鉄ホメオスタシスのエピジェネティック制御により葉の老化を遅らせる
Histone variant HTB4 delays leaf senescence by epigenetic control of Ib bHLH transcription factor-mediated iron homeostasis
Qi Yang1* , Ting Wang1*, Jie Cao1*, Hou-Ling Wang1 , Shuya Tan1, Yuan Zhang1, Sanghoon Park2, Hyunsoo Park2, Hye Ryun Woo2,3 , Xiaojuan Li1 , Xinli Xia1 , Hongwei Guo4 and Zhonghai Li1
New Phytologist (2023)
doi: 10.1111/nph.19008
(概要)
葉の老化は、複数の内的因子と栄養欠乏を含む多様な環境ストレスによって制御される秩序だったプロセスである。ヒストン変異体は植物の成長と発達の制御に関与している。
しかし、葉の老化におけるヒストン変異体の機能やその制御機構については、まだ不明な点が多い。
ここでは、H2Bヒストン変異体HTB4が葉の老化の負の制御因子として機能することを明らかにした。HTB4の機能欠損は、葉の老化の初期表現型を引き起こし、機能相補性によって回復した。
RNA-seq解析により、いくつかのIbサブグループ,
塩基性ヘリックス-ループ-ヘリックス(bHLH)が鉄(Fe)のホメオスタシスに関与する転写因子(TF)であることが明らかになった。
htb4変異体では、bHLH038、bHLH039、bHLH100、bHLH101などの鉄(Fe)恒常性発現が抑制され、FERRIC REDUCTION OXIDASE 2 と IRON-REGULATED TRANSPORTER (IRT1) の発現を低下させた。 クロマチン免疫沈降-定量ポリメラーゼ連鎖反応解析により
HTB4はIb bHLH TFのプロモーター領域に結合し、転写開始点近傍の活性マークH3K4me3の濃縮を促進することにより、その発現を増強することが明らかになった
さらに、Ib bHLH TFまたはIRT1を過剰発現させると、htb4変異体の早期老化表現型が抑制された。
我々の研究により、HTB4-bHLH TFs-FRO2/IRT1-Feホメオスタシスという、 葉の老化の発症と進行を制御するシグナル伝達経路を確立した。
(はじめに)
葉の老化は、秋の壮大な景観を作り出す一般的な自然現象である。
葉の老化の始まりは 落葉植物では、緑色から黄色、橙色、赤色へと色が変化する(Keskitalo et al. 2005).
葉の老化では、核酸、脂質、タンパク質などの高分子が分解され、代謝され、 放出された栄養分は、新芽や果実、種子などの活発な生長器官に移動するか、あるいは、 次の生育期に向けて貯蔵される(Lim et al., 2007; Guo & Gan, 2014)。
したがって、 葉の老化は植物のフィットネスにとって重要であり、育種家にとって非常に興味深い形質である。なぜなら、早すぎる老化は作物の収量、ポストハーベスト 貯蔵、品質に悪影響を及ぼすからである(Guo & Gan, 2014)。
葉の老化の重要な制御成分とその根底にある分子機構を明らかにすることは 農業や森林における分子育種に貴重な情報を提供する (Gepstein, 2004; Wang et al., 2021)。
葉の老化は、何千もの老化関連遺伝子(SAG)によって制御される高度に協調的なプロセスであり、これらの遺伝子の転写レベルは葉の老化に伴って変化する(Gan & Amasino, 1997; Lim et al.)
葉の老化を制御するいくつかの重要な遺伝子は、老化の表現型が変化した突然変異体の前方遺伝学的スクリーニングによって同定された(Lim et al.2007)
ゲノム情報の利用可能性とゲノム編集技術の登場により、葉の老化に関する研究はますます飛躍的に進展している。
逆遺伝学的アプローチにより、数十の植物種における数百のSAGの機能評価が可能になった(Woo et al.)
マルチオミクス研究により、葉の老化は非常に複雑な遺伝的プログラムであり、主に転写、転写後、翻訳、翻訳後の制御など、複数の層の制御によって細かく制御されていることが明らかになった(Wooら、2013、2019;Caoら、2022)。
その中でも、転写調節は 葉の老化の開始と進行を制御する上で重要な役割を果たしている(Kimら、2016)。
転写因子(TF)は、葉の老化の間、SAGの発現の劇的な変化を促進する中心的な制御要素として働く(Kim et al.)
シロイヌナズナでは、NAC、WRKY、MYB、bHLH、bZIPファミリーのメンバーなど、数多くのTFが葉の老化制御に関与していることが判明している(Limら、2007;Balazadehら、2008;Wooら、2019;Liら、2020)。
ヒストンメチル化、ヒストンアセチル化、DNAメチル化などのエピジェネティック修飾の変動は、葉の老化プロセスに影響を与える。
例えば、JUMONJI DOMAIN-CONTAINING PROTEIN 16 (JMJ16)はヒストンH3リジン4トリメチル化(H3K4me3)脱メチル化酵素であり、葉の老化を遅らせる(Liu et al.
SAGのサブセットの転写リプログラミングを通じて葉の老化を促進する(Wang et al.) DEMETER様DNA脱メチル化酵素遺伝子DML3は、主に葉の老化期に発現する。
DML3の変異はDNAの高メチル化をもたらし、いくつかのSAGの発現を阻害することで、葉の老化を遅らせる(Yuan et al.)
老化の開始と進行は、老化プロセスに統合されたさまざまな内部シグナルや環境的な合図によっても影響を受ける(Guo & Gan, 2005; Limet al., 2007)。
干ばつ、極端な温度、栄養不足などの生物学的ストレス要因や、病原体感染などの生物学的要因は、葉の老化プロセスを加速させる可能性がある。
窒素は植物にとって重要な大栄養素であり、その欠乏は葉の老化を早める。
NITROGEN LIMITATION ADAPTATION (NLA)は、窒素飢餓によって誘導されるRING型ユビキチンE3リガーゼであるNLAは、ORESARA 1(ORE1)と相互作用し、ポリユビキチン化を通じてその安定性を制御している。
ORE1の機能喪失は、窒素欠乏が引き金となる葉の老化を遅らせるが、nla変異体 は窒素が不足すると急速に老化が進行する。
植物の必須微量栄養素である鉄(Fe)は、光合成、呼吸、ホルモン生合成、形態形成など、生長および発育プロセスにおいて重要な役割を果たしている。ホルモンの生合成、形態形成、細胞酵素反応などである。
鉄欠乏は早期の葉の老化を誘発し、作物の生産性と食用穀物の栄養品質を制限する(Higuchi et al.)
しかし、葉の老化の開始と進行を制御するために、様々な制御因子を調整する基本的なメカニズムは、まだ解明されていない。
今回我々は、ヒストン変異体HTB4が、葉の老化の新たな負の制御因子として機能することを発見した。
HTB4の機能欠損は葉の老化を早めるが、HTB4本来のプロモーターで発現させることで回復した。
さらに、HTB4の変異は、鉄欠乏に応答するbHLH TFをコードする遺伝子の発現に影響を与え、その結果、内因性鉄レベルが低下した。以上の結果から、HTB4はbHLH TFを介した鉄ホメオスを制御していることが示唆された。制御していることが示唆された。
これらの結果は葉の老化に関する新たな制御パラダイムを提供するものである。
以下図の説明
(図1)
HTB4はシロイヌナズナの葉の老化の負の制御因子である。
(a) 24日齢のCol-0、htb4およびHTB4comp植物の老化表現型。
HTB4compの2つの独立した系統、#1と#3を調べた。バーは2cm。
(b) Col-0、htb4およびHTB4comp植物のロゼット第3葉および第4葉の表現型。代表的な葉を示す。棒は1cm。
(c) クロロフィル含量と
(d) (b)の葉におけるFv/Fm比。
(e) 20日齢の葉におけるSAG12の発現。
(図2) HTB4はシロイヌナズナの鉄ホメオスタシスに関与している。
(a)(+Fe)または鉄欠乏(Fe)条件下でのHTB4pro:GUS/Col-0トランスジェニック植物の組織化学的解析。
½ MS培地で7日間生育させた苗を、半分の強さのMS液体培地(1/2MS)、0.1mM Fe EDTA(+Fe)、または0.05mM フェロジン(Fe)添加Fe欠乏培地にそれぞれ3時間移植し、GUS染色に用いた。
(b) (a)と同様にして、植物体を鉄過剰(+Fe)または鉄欠乏(Fe)条件下でHTB4pro:GUS活性を測定した。
GUS活性は計算し、1分間に1lgのタンパク質あたりの4-メチルウンベリフェロン(4-MU)のピコモルとして表した。
(c) 10日齢のCol-0、htb4およびHTB4comp植物の苗における鉄含有量。
苗の元素濃度は誘導結合プラズマ発光分光分析(ICP-OES)で測定した。
(d) 定性ポリメラーゼ連鎖反応(RT-qPCR)による FERRIC REDUCTION OXIDASE 2 (FRO2)発現の解析。
植物を MS 培地で 10 日間生育させた後、鉄を添加した培地または無添加の培地に 移し、さらに 4 日間生育させた。FRO2の相対量はACT2に対して正規化した。
(e) Col-0、htb4およびHTB4comp植物における鉄キレート還元酵素活性の測定。
植物はMS培地で10日間生育させ、さらに4日間Fe添加または無添加の培地に移植した。GUS、b-グルクロニダーゼ。
(図6 )
葉の老化制御におけるHTB4-bHLH TFs-FRO2/IRT1-Feシグナル伝達カスケードを示すモデルの提案。
われわれの研究および過去の報告に基づいて、葉の老化の発症と進行を制御するHTB4-bHLH TFs-FRO2/IRT1-Feホメオスタシスというシグナル伝達経路を確立した。
ヒストン変異体HTB4は、bHLH038、bHLH039、bHLH100、bHLH101を含むIbサブグループのbHLH TFsのクロマチンに結合し、遺伝子発現を活性化するH3K4me3修飾を仲介する。
Ib bHLH TFsはFITとヘテロダイマーを形成することにより、IRT1とFRO2の転写を活性化し、鉄の取り込みと恒常性を高め、葉の老化を遅らせる。
さらに、Ib bHLH TFは HTB4の発現も制御している。HTB4は葉の老化の機能的制御因子であり、その転写は年齢と鉄欠乏によって制御される。HTB4は、加齢や環境因子によって誘導される葉の老化を抑制する収束点の1つとして機能している可能性が示唆された。
矢印は活性化、鈍端矢印は抑制を表す。 FRO2、FERRIC REDUCTION OXIDASE 2; Fe、鉄; TFs、転写因子。
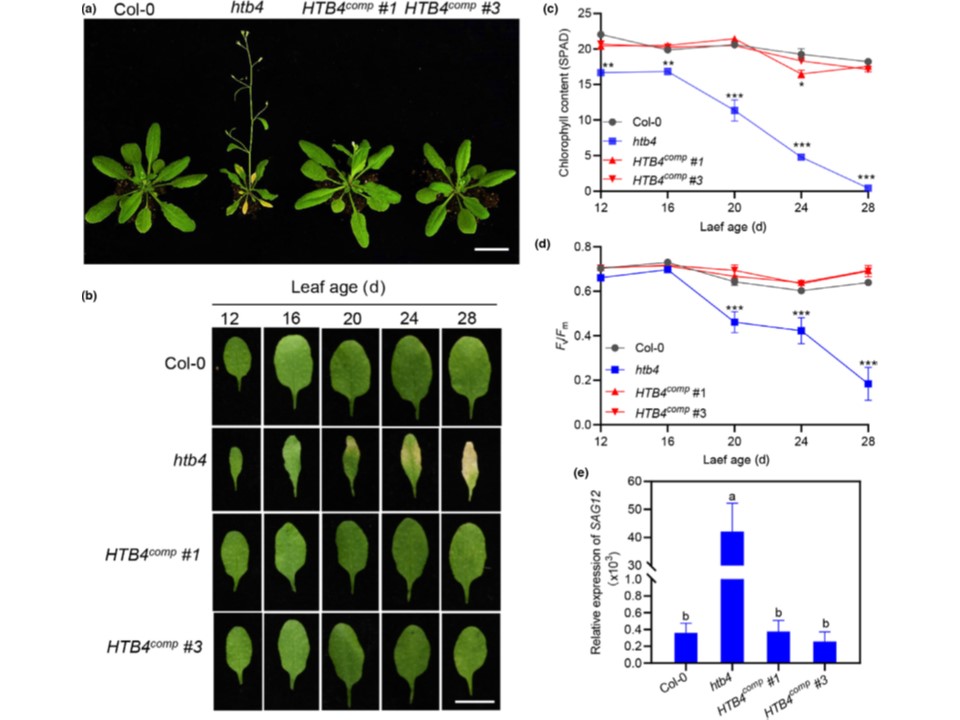
図1
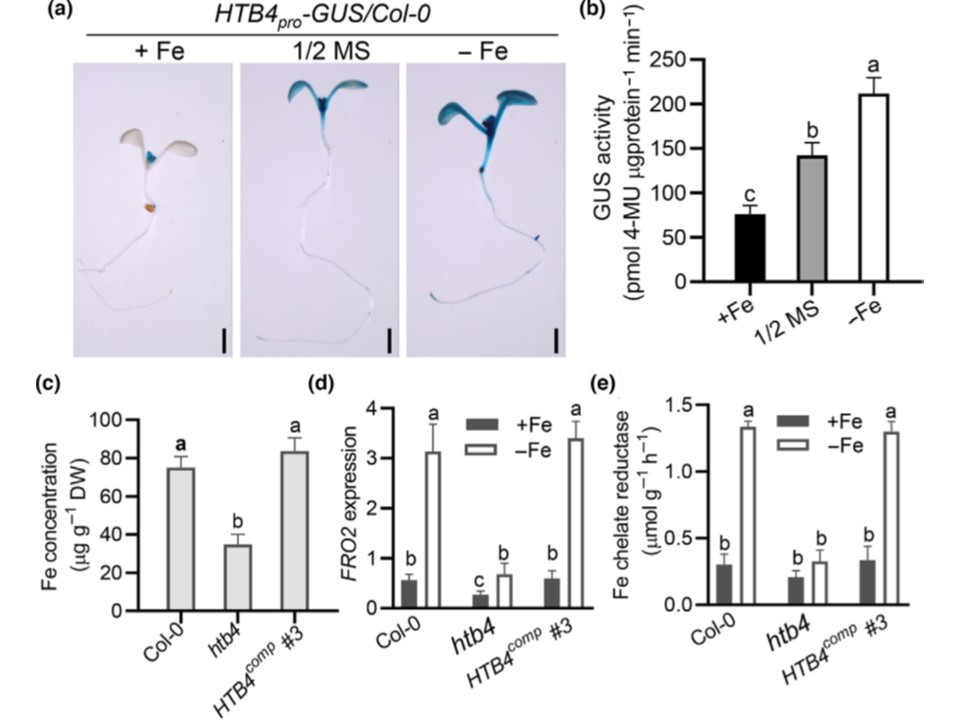
図2
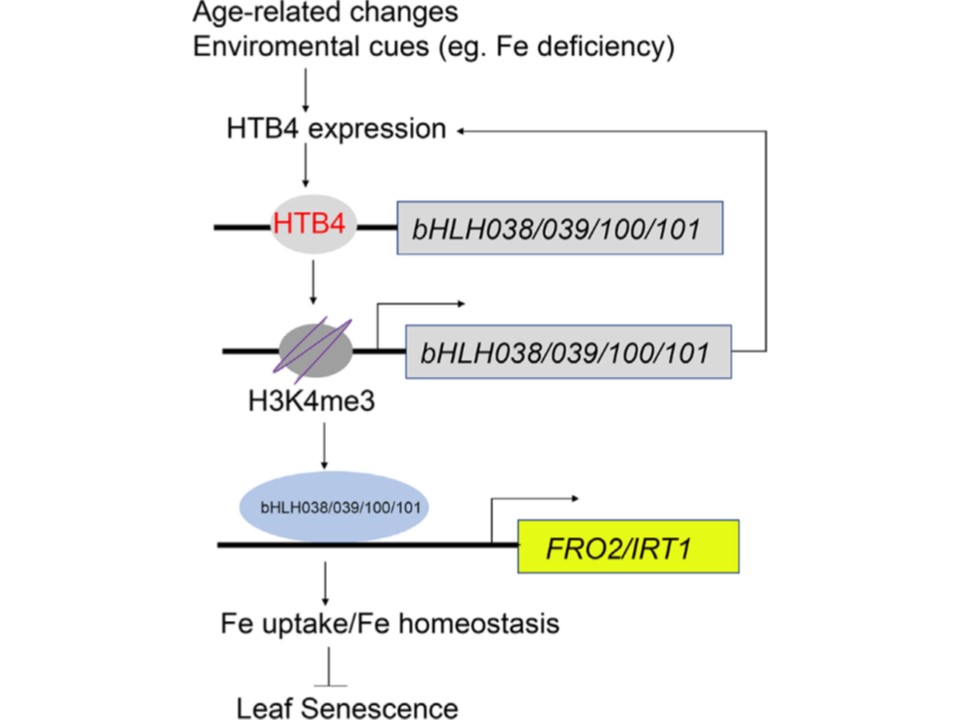
図6