トランスクリプトーム解析と機能解析により、ジャガイモウイルスY感染に対するタバコの鉄媒介抵抗性の分子機構が明らかになった
鉄の葉面散布がポテトウイルスの初期感染抑止に効果があるという報告である。小生にはちょっと信じがたいが、感染症の専門でないので、批評できない。
Translate transcriptomic and functional analyses reveal the molecular mechanisms underlying Fe-mediated tobacco resistance to potato virus Y infection from English to Japanese
トランスクリプトーム解析と機能解析により、ジャガイモウイルスY感染に対するタバコの鉄媒介抵抗性の分子機構が明らかになった
徐春桃1,2†、郭慧艳1†、李睿1、蓝新宇1、張永輝2、谢強2、朱迪3、穆青3、王志平1、安孟楠1、夏子豪1*、呉元華1* 1 農業大学植物保護学院、遼寧省植物病理学重点実験室、沈陽、中国 2 四川省タバコ会社泸州市会社、泸州、中国 3 貴州省黔西南州タバコ会社、興義市、中国
Front. Plant Sci. 14:1163679. doi: 10.3389/fpls.2023.1163679
要旨
ポテトウイルスY(PVY)は主にナス科作物に感染し、収量と品質に大きな損失をもたらします。鉄(Fe)は植物のさまざまな生物学的過程に関与していますが、PVY感染に対する抵抗性におけるその役割は報告されていません。本研究では、鉄の葉面散布がPVYの初期感染を効果的に抑制することができることを示し、PVY感染したニコチアナ・タバクムでその作用機序を調べるために、フルレングス転写組織学とイルミナRNAシーケンシングを行いました。その結果、18,074個の代替スプライシング変異体、3,654個の融合転写産物、3,086個の長鎖非コードRNAおよび14,403個の発現量変動遺伝子(DEGs)が同定されました。特に、鉄処理は接種後1日目(dpi)にリン脂質加水分解、リン脂質シグナル、細胞壁生合成、転写因子(TFs)および光系I組成に関連するDEGsの発現レベルを低下させる一方で、光合成電子伝達系(PETC)に関与するDEGsの発現レベルを上昇させました。接種後3日目では、光系II組成、PETC、分子シャペロン、タンパク質分解および一部のTFsに関連するDEGsが上昇しましたが、光収集系、リン脂質加水分解および細胞壁生合成に関連するDEGsは低下しました。接種後9日目では、鉄処理はPVY感染への抵抗性や転写プロファイルにほとんど影響を与えませんでした。これらの潜在的に重要なDEGsの機能解析はその後ウイルス誘導遺伝子サイレンシング法を用いて行われました。
論議(一部省略)
本研究では、PVY感染によるN. tabacum遺伝子発現への影響と、PVY感染に対するFe施用の分子基盤を、感染1、3、9日後のPacBio SMRTおよびIllumina RNAシーケンスによってより深く理解した。
トランスクリプトーム解析により、PVY感染は主に光合成や、脂質代謝やタンパク質処理などのほとんどの生物学的プロセスに関連する遺伝子の発現レベルに影響を与えることがわかった。
この結果から、鉄の施用は、主に光合成、脂質代謝、タンパク質処理、細胞壁生合成、病害抵抗性に関連する転写因子群(TFs)などの遺伝子の発現レベルを感染初期に調節することにより、PVY感染に対するタバコの抵抗性を向上させることができると考えられた。
転写因子は植物遺伝子の発現を制御し、ストレスへの応答において重要な役割を果たすことができる。
本研究では、植物免疫において極めて重要な役割を果たすことが報告されている6つのファミリー(NAC、WRKY、MYB、bHLH、GATA、TCP)から、合計227個の発現量の異なる転写因子をRNA-Seqにより同定した。
感染1日後ではNtbHLH30とNtGATA8がダウンレギュレートされ、3日後ではPVY + Fe vs. PVY + HでNtNAC2、NtMYB4、NtWRKY22がアップレギュレートされていることがわかった。以前には、N. benthamianaにおけるNbMYB4Lの過剰発現がTMVに対する有意な抵抗性を誘導することが実証されている。
Paeonia lactifloraの葉におけるPlWRKY65は、病原体関連(PR)遺伝子の発現を誘導し、ジャスモン酸(JA)含量を増加させることにより、A. tenuissimaに対する抵抗性を高めることが実証されている。
カブ・クリンクルウイルス(TCV)のCPタンパク質は、シロイヌナズナのNAC転写因子であるTIPと相互作用し、サリチル酸経路を阻害してウイルス感染を促進することができる。
従って、鉄の施用は、これらの上記転写因子の発現を制御することにより、病害抵抗性遺伝子の発現を誘導し、その結果、PVY感染に対するタバコの抵抗性を高めることができると仮定された。
また、NbWRKY26(NtWRKY22のホモログ遺伝子)をサイレンシングすると、N. benthamianaにおけるPVYの蓄積が促進されたことから、NbWRKY26がPVY感染に対する抵抗性に関与する可能性が示された。
光合成は植物の生命活動の基礎であり、あらゆる生命活動にエネルギーを供給する。
PVYに感染したジャガイモのプロテオーム解析では、ウイルス感染がクロロフィルa-b結合タンパク質の発現、および植物の光合成に大きな影響を与えることが示されている。
本研究では、PVY感染9日後で多種多様な光合成関連遺伝子の発現量が低下していることを確認した。
感染1日後では、PVY+Fe対PVY+HでNtCyt b6、NtPSI-A2、NtPsaG、NtPcが増加し、感染3日後ではNtCabsがダウンレギュレーションしていることがわかった。
さらに、NbCab-6AのPVY感染抵抗性に対する役割をTRVベースのVIGSで検証した結果、NbCab-6Aをサイレンシングした植物が高いPVY抵抗性を示すことが明らかになった。
ストロベリーベインバンディングウイルス(SVBV)がコードするP1は、光収穫複合体IIタイプ1ライクのクロロフィルa/b結合タンパク質(LHC II-1L)と相互作用でき、LHC II-1Lの過剰発現はSVBV感染を促進することが報告されている。
したがって、NbCab-6AはPVYの感染を促進したのは、おそらくPVYがコードするタンパク質と直接または間接的に相互作用するためであり、さらなる研究が必要である。
植物には多くの種類の脂質が存在し、それらは主に膜構造組成に関係している。
脂質代謝における複数の中間体(すなわちPLD、PI-PLC)は、ジャスモン酸(JA)やサルチル酸(SA)シグナルの制御や宿主のPAMP-triggered immunity(PTI)応答の誘導など、様々なストレス応答に関与することが報告されている。
本研究では、PVY + FeとPVY + Hの比較において、1日後でのNtPLD、NtPIPPase、NtnsLTP、3日後でのNtPLA、NtPLD、NtFADの集積が低下していた。ジャガイモでStLTP6を過剰発現させると、RNAサイレンシング経路に関わる遺伝子の発現が抑制され、PVYおよびポテトウイルスS(PVS)の感染が促進されることが研究で示されている。
StLTP10を過剰発現させると、病害抵抗性遺伝子の発現を改善し、酸化ストレスを軽減してPhytophthora infestansに対する抵抗性を高める。
脂肪酸脱変性(FAD)は不飽和脂肪酸(UFA)の生成を触媒し、JA、SA、テルペンなどの各種ホルモンの生合成の前駆体となる。
シロイヌナズナのfad3fad7fad8三重変異体は、SA合成が障害され、その結果、オミクシスに対する感受性が向上する。
本研究では、N. benthamianaにおいてNbnsLTPまたはNbFAD3をサイレンシングすると、PVY感染に対する感受性が高まることを見いだした(図9)。
これらの結果から、NbFADとNbnsLTPはPVY感染に対する抵抗性に重要な役割を担っており、ホルモンシグナル伝達や酸化ストレス応答に対する制御と関連している可能性が示唆された。
植物におけるタンパク質の折り畳みと修飾は、主に小胞体で行われる。
HSPやDnaJなどの分子シャペロンは、植物の小胞体におけるタンパク質の折り畳みを補助している。
Calreticulin(CRT)は小胞体内腔の可溶性タンパク質で、他の分子シャペロンを呼び寄せてグリコシル化タンパク質の折り畳みを促進する。E3ユビキチンリガーゼは主にミスフォールドしたタンパク質を分解する。
本研究では、PVY + HとP + Hの比較において、NtHSP90、NtHSP20、NtDnaJ、NtCRTが感染9日後でアップレギュレートされることが示された。
興味深いことに、NtHSP90、NtHSP20、NtE3もPVY + Fe vs. PVY + Hの感染3日後で発現が上昇した。
VIGSアッセイにより、NbHSP90をサイレンシングした植物では、PVY RNAとCPタンパク質の蓄積量が対照植物と比較して高いことを見いだした。
これまで、N. benthamianaでNbHSP90をサイレンシングすると、PR遺伝子の発現量が低下し、ポテトウイルスX(PVX)、P. syringae、TMVの感染を促進することが多くの研究で示されてきた。
一方、最近の研究では、N. benthamianaでジャガイモStHSP90.5の相同遺伝子をサイレンシングすると、PVYの蓄積を抑え、防御関連遺伝子発現を誘導することが示されている。
N. benthamianaのNbHSP90遺伝子をサイレンシングすると、PR遺伝子の発現を誘導することでRalstonia solanacearumの感染を抑制することができたと報告されている。
N. benthamianaの異なるNbHSP90は多様な機能を有している可能性があり、PVYに対する抗ウイルス活性における役割についてさらに検討する必要があると考えた。
植物細胞壁のリグニン化とカロース沈着は、病原体の感染を防ぎ、シグナル分子を伝達することで抵抗性応答を誘導する。
本研究では、PVY + H vs. P + Hにおいて、木質化およびカロース沈着に関与するNtCESA、NtLaccase、NtCaISが感染9日後でダウンレギュレートしていることを明らかにした。
NtCESA、NtLaccase、NtCaISの発現レベルは、PVY + FeとPVY + Hを比較すると、感染一日後と3日後ですべて低下していた。
セルロース合成酵素は、植物の細胞壁形成時のセルロース合成に関与している。
Botrytis cinereaがシロイヌナズナに感染すると、セルロース合成酵素遺伝子の発現量が低下し、耐病性が向上することから、シグナル伝達を通じて免疫反応が活性化されると考えられている。
植物ラッカーゼはリグニン合成に関与することで細胞壁合成を制御している。
綿花のGhLac1遺伝子の発現を抑制することで、JAや二次代謝物の蓄積をもたらし、バーティシリウム・ダリアに対する抵抗性を向上させることができる。
カロースの沈着は植物の自然免疫反応の一部であり、カロースはカロースシンターゼによって合成される。
柑橘類のCalS1遺伝子をサイレンシングすると、Xanthomonas citri subsp. citriの感染が促進された。
カロースの沈着はアブシジン酸含量と密接に関係しており、シロイヌナズナではアブシジン酸含量が低いと抑制される。
以上のことから、FeはNtCESAおよびNtLaccase遺伝子の発現を阻害することにより、細胞壁シグナルの細胞内流動を促進し、免疫シグナル伝達を活性化し、病害抵抗性に関連する二次代謝産物の蓄積を誘導することにより、おそらくPVY感染に対する宿主抵抗性を高めると推察された。
図1
外因性Feの葉面散布は、タバコ植物におけるPVY感染を緩和した。
(A) 4つの処理における7日目での植物全体、葉および茎の症状。
(B) 4つの処理における9日目の植物全体、葉および茎の症状。
(C) 1、3、5、7、9日目のRT-qPCRで測定したPVY + HおよびPVY + Feグループのタバコ葉におけるPVYゲノムRNAの蓄積量。アスタリスクは、コントロールと比較して統計的に有意な差を示す(Student's t-test): *p < 0.05, **p < 0.01.
(D)1、3、5、7、および9日目でウェスタンブロットアッセイにより決定したPVY+HおよびPVY+Feグループのタバコ葉におけるPVY CPタンパク質の蓄積量。
図9
N. benthamianaにおけるTRVベースのVIGSアッセイによるPVY感染に対する抵抗性の5つの相同遺伝子の機能解析。
(A) PVY感染後の異なる遺伝子サイレンサーN. benthamiana植物における病徴。
(B) (A)の白抜き枠で示した上葉のクローズアップ写真。
(C) 異なる遺伝子サイレンシングを施したN. benthamianaにおけるRT-qPCRで測定したPVYゲノムRNAの蓄積量。アスタリスクは、両側t検定により決定された処理間の統計的差異を示す(*, P < 0.05; **, P < 0.01).
(D) N. benthamianaの上葉におけるPVY CPタンパク質の発現レベル。CP、コートタンパク質、CBB、クマシーブリリアントブルー。
(E) RT-qPCRにより決定された標的遺伝子のサイレンシング効率(Silencing Efficiency)。アスタリスクは、両側t検定により決定された処理間の統計的差異を示す(*, P < 0.05; **, P < 0.01).
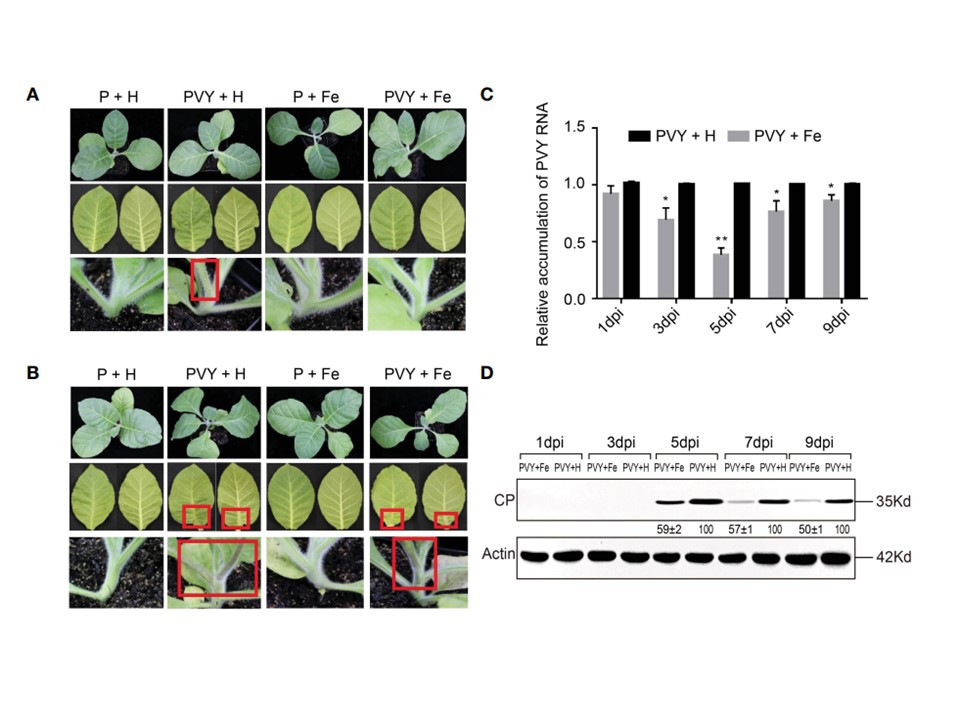
図1
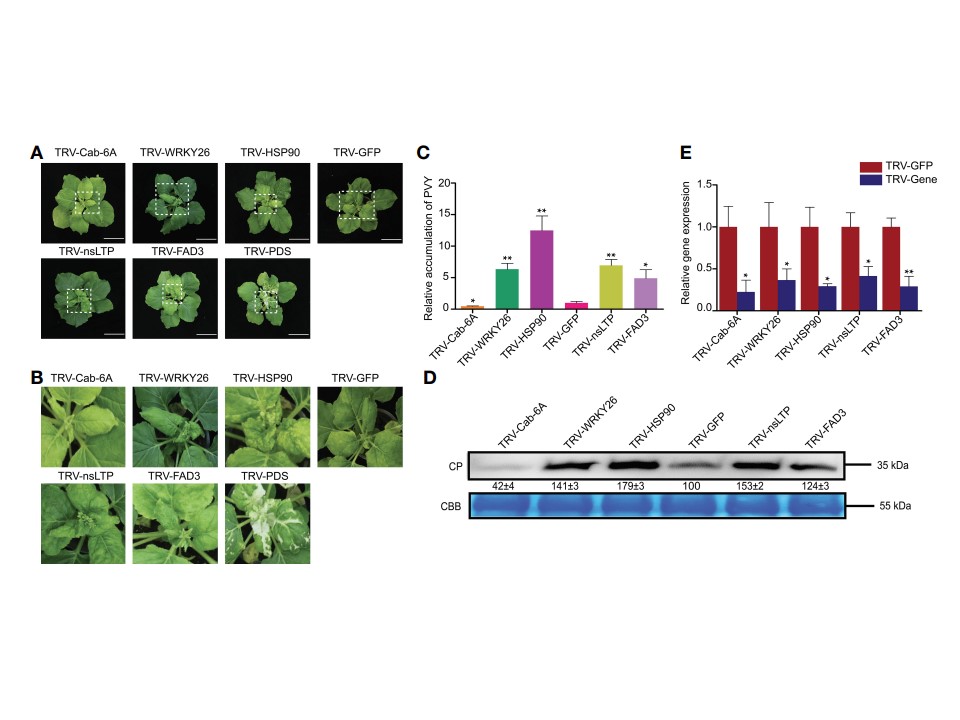
図9