総説:鉄貯蔵プールを超えて:ストレス下における植物アポプラスト鉄の機能
アポプラストの鉄の挙動や役割についての研究はそれほど多くない上に体系的に研究されてこなかった。中国の研究者グループは、その隙間を狙って体系化を試みようとしている。
総説:鉄貯蔵プールを超えて:ストレス下における植物アポプラスト鉄の機能
Beyond iron-storage pool: functions of plant apoplastic iron during stress
Xing Xing Liu , Xiao Fang Zhu , Da Wei Xue , Shao Jian Zheng , and Chong Wei Jin
Trends in Plant Science, Month 2023, https://doi.org/10.1016/j.tplants.2023.03.007
(概要)
鉄(Fe)は植物にとって必須微量栄養素であり、アポプラストへの貯蔵は重要なFeプールとなる。
植物は、このアポプラスト鉄プールを再利用して、鉄欠乏に適応するための様々な戦略を開発してきた。
さらに、アンモニウムストレスやリン酸欠乏症や病原体からの攻撃など、他のストレスに対する植物の適応には、アポプラストFeの動的変化が重要であることを示す証拠が増えている。
このレビューでは、ストレスの刺激に対する植物の行動変化におけるアポプラスト鉄の関連性について議論し、精査する。
我々は主に、ストレスシグナリングネットワークにおけるアポプラスティックFeの作用と下流事象を調節する関連成分に焦点を当てる。
図2. 鉄欠乏条件下でのアポFeの再利用。根では、鉄欠乏によりフェノール排出トランスポーター(AtPDR9やOSPEZ2など)の発現が上昇し、固定化されたアポプラスト鉄(III)をキレートするためにフェノールを分泌する。
非イネ科植物では、Fe(III)-フェノールキレートは、その後Fe(II)トランスポーター(例えば、AtIRT1)によって取り込まれるための鉄キレート還元酵素(例えば、AtFRO2)によりFe2+に還元されます。
イネ科植物では、MA排出トランスポーター(例えばOsTOM1)によって分泌されるムギネ酸(MA)ファミリーファイトシデロフォアも固定化されたアポプラスト鉄(III)をキレートし、MA-Fe(III)キレートは黄色ストライプ様トランスポーター(YSL)によってシンプラスト内に輸送される可能性があります。
中心柱では、クエン酸排出トランスポーター(AtFRD3やOsFRDL1など)が分泌するクエン酸やフェノール排出トランスポーター(OsPEZ1/2など)が分泌するフェノールが、固定化したアポプラスト鉄(III)のキレートに関与している可能性が考えられた。
また、グリコシルトランスフェラーゼAtCdiによるラムノガラクツロナン-II(RG-II)のガラクトシル化など、細胞壁のカチオン結合部位の修飾も、根と葉の両方で固定化アポプラストFe(III)の放出を調節している。
葉の中葉細胞のNO3 - トランスポーターNRT1.1は、鉄欠乏によりダウンレギュレーションされ、H+共役NO3 - 取り込みを減少させ、アポプラスト内のH+を十分に維持し固定化Fe(III)を放出させる。
また、鉄欠乏により葉のアポプラスト中のクエン酸やリンゴ酸が増加することも、固定化された鉄(III)を放出するのに有利であると考えられる。
鉄キレート還元酵素(AtFRO6など)とアスコルビン酸は、葉緑体細胞が鉄を取り込むために、放出された葉緑体の鉄(III)を還元すると考えられる。根の導管から輸送されるクエン酸は、キレート化された導管のアポ-Feとしての役割を果たすことができる。
フェロポーティン(例えば、AtFPN2)[122]によって根の導管にFe2+がロードされ、クエン酸がFe(III)をキレートするのに好まれるため、根の師管と葉脈の両方のアポプラストでFeの固定化を避けるためにフェロキシダーゼ(例えば、AtLPR1およびAtLPR2)が必要である。
図3.
Apo-FeはアラビドプシスのNH4 +ストレスおよびP欠乏に対する根の発達を再形成する。
(A) NH4 + ストレス下での根の成長阻害のメカニズム。
NH4 + ストレスは、細胞壁フェロキシダーゼ LPR2 に依存して根の葉茎にアポ-Fe を過剰蓄積させ、活性酸素種(ROS)バーストを引き起こしてカロース沈着を誘発する。
その結果、NH4 +ストレスは、スクロースの成長領域への配分が不十分であるため、根の成長を抑制する。
AtPDX1.1を介したビタミンB6(VB6)の合成は、過剰な活性酸素に対抗し、アポFe依存的な成長阻害から根を保護する。
(B)P欠乏(-P)下での根の成長阻害のメカニズム。
-Pは、主に細胞壁フェロキシダーゼLPR1を介して、幹細胞ニッチ(SCN)および/または伸長帯(EZ)皮質細胞のアポプラストに鉄を蓄積させ、ALMT1を介したリンゴ酸排出により促進される。
LPR1およびALMT1の転写を活性化するには、それぞれHY5およびSTOP1が必要である。
その後、アポ鉄の過剰蓄積により活性酸素が蓄積され、ペルオキシダーゼ(PRX)に依存した細胞壁の硬化が進み、カロース沈着によるプラスモデスマータ(PD)の閉鎖によりSHRの細胞間移動が制限され、根の成長が阻害される。
また、いくつかのリプレッサーも-P下でアポ鉄の蓄積を負に制御している。BZR1とBRMを介したHDC1-HDA6複合体のLPR1遺伝子座への動員はともにLPR1転写を抑制するが、後者の負の制御は-Pにより減弱する。
PDR2およびALS3-STAR1モジュールもまた、アポ-Feを負に制御する は、未知のメカニズムでLPR1およびSTOP1の作用をそれぞれ制限することにより、-P下での蓄積を抑制する。
図4.
病原体の侵入に対するアポプラストFeに関連した植物の防御機構。
左のパネルは、アポ-Feダイナミクスに関連するFe保留機構を描いている。
植物のEDS1-RPS4複合体は、細菌性病原体のエフェクターであるAvrRps4を認識して、BTSによるbHLH105とbHLH115の分解の妨害を防ぎ、葉にアポFeが蓄積して病原体の増殖が制限される。
さらに、植物はPDF1.1をアポプラストに分泌して、利用可能なPDF1.1-Fe複合体を形成するため、それ自体が植物の細胞内Fe獲得を阻害し、Fe欠乏応答性のエチレンバーストを誘発して細菌病原体感染を抑制することもできる。
右図はアポ-Feの過剰蓄積機構を示す。
菌類に感染すると、植物の細胞質鉄は、鉄を含んだ小胞体状の小胞体が運ばれて、感染由来のアポプラストに集められる。感染由来のアポプラストは、鉄を結合する植物細胞壁の高分子がないため、鉄を活性型で提示して防御的酸化バーストを誘発することができる。
略号 ROS, 活性酸素種.
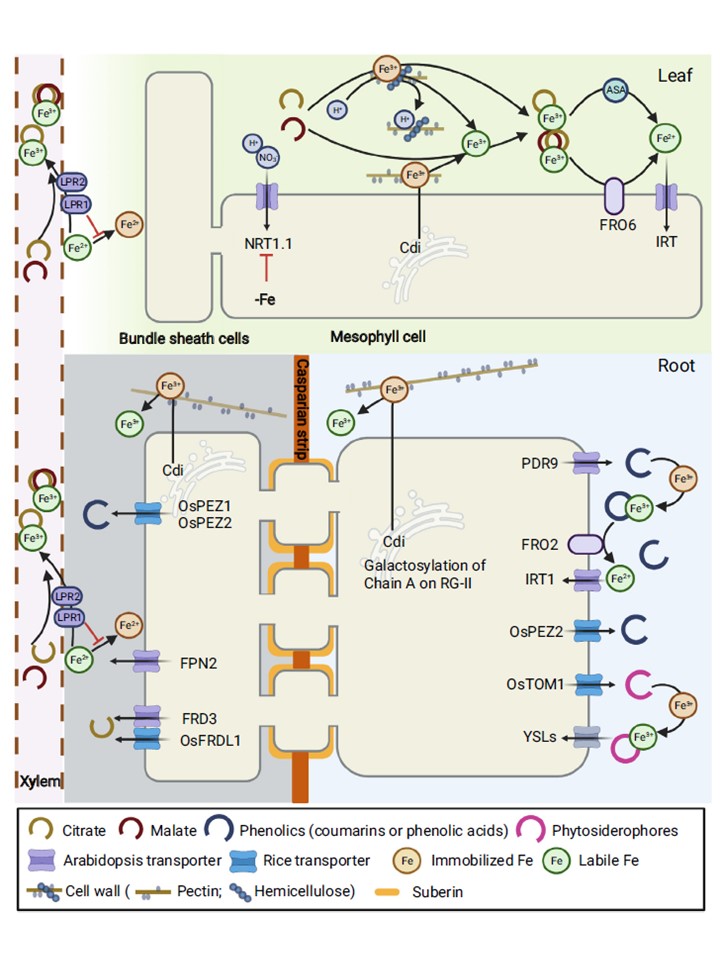
図2
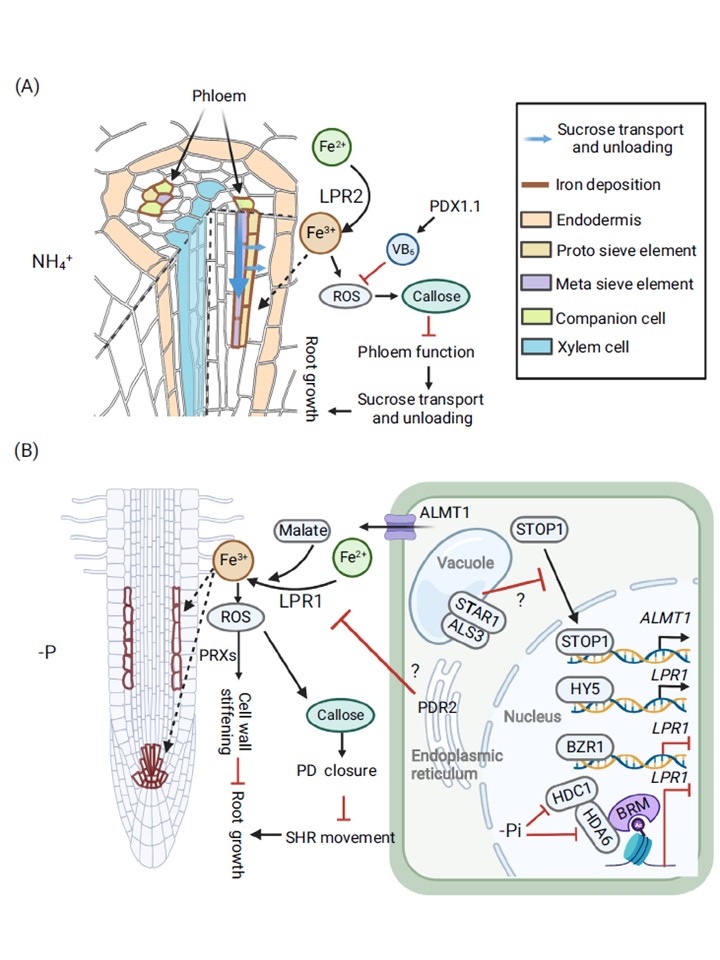
図3
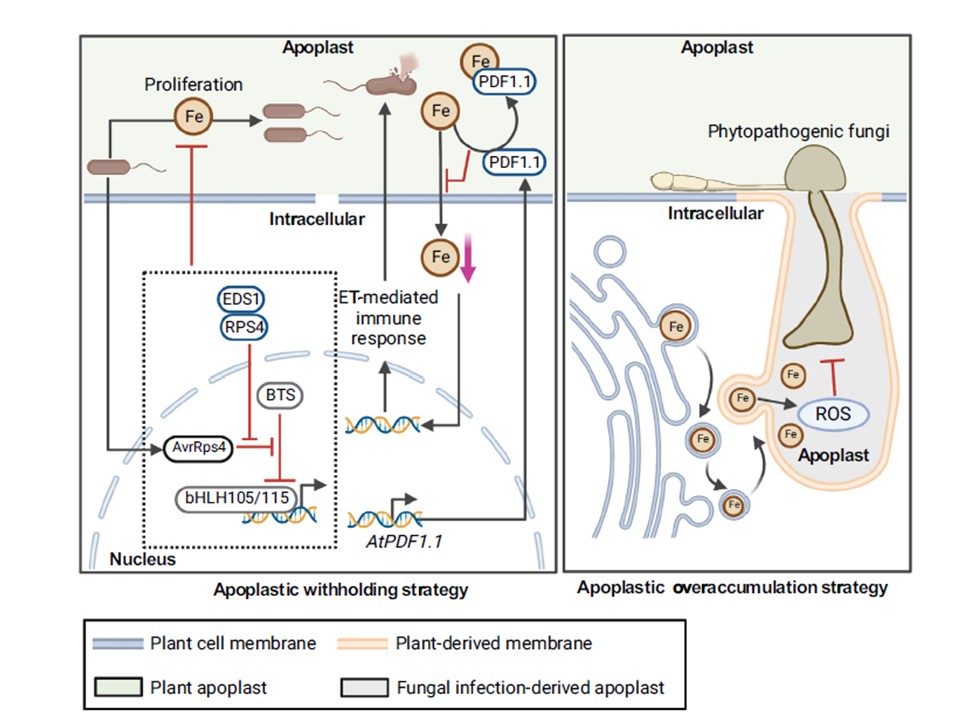
図4