Pseudomonas aeruginosa乮椢擽嬠乯偑揝偵傾僋僙僗偡傞暋悢偺愴棯
偙偙偱偼彫惗偑偙偺榑暥偱採帵偝傟偰偄傞懡偔偺偦傟偧傟偺僔僨儘僼僅傾偺庢傝崬傒婡峔儌僨儖偺偆偪丄怉暔偵傕懡彮娭學偡傞偲巚傢傟傞乽僋僄儞巁揝乿偲乽僿儉揝乿偺媧廂婡峔偵偮偄偰偺傒徯夘偡傞丅
Pseudomonas aeruginosa乮椢擽嬠乯偑揝偵傾僋僙僗偡傞暋悢偺愴棯
Pseudomonas aeruginosa and its multiple strategies to
access iron
僀僓儀儖丒J丒僔儍儖僋1,2乥僋僃儞僥傿儞丒儁儘乕1,2
乮奣梫乯
Pseudomonas aeruginosa乮椢擽嬠乯偼丄懡偔偺帺慠娐嫬偍傛傃恖岺娐嫬偵懚嵼偡傞儐價僉僞僗側嵶嬠偱偁傞丅
傑偨丄怉暔丄摦暔丄僸僩偺昦尨懱偱傕偁傞丅
傎偲傫偳偺惗暔摨條偵丄揝偼椢擽嬠偺憹怋偵晄壜寚側塰梴慺偱偁傞丅
椢擽嬠偼丄懡條側帺慠娐嫬偍傛傃廻庡娐嫬偵偍偄偰惗偒巆傞偨傔偵丄揝偵傾僋僙僗偟丄偦偺儂儊僆僗僞僔僗傪堐帩偡傞偨傔偺暋嶨側僔僗僥儉傪恑壔偝偣偰偒偨丅
P. aeruginosa偼丄揝傪棙梡偡傞偨傔偵丄2庬椶偺僔僨儘僼僅傾乮僺僆儀儖僕儞丄僺僆僔僃儕儞乯傪惗嶻偡傞偙偲偑偱偒傞丅傑偨丄懠偺嵶嬠偑惗嶻偡傞條乆側僔僨儘僼僅傾乮儅僀僐僶僋僠儞丄僄儞僥儘僶僋僠儞丄僼僃儕僆僉僒儈儞丄僼僃儕僋儘儉丄價僽儕僆僶僋僠儞丄僄傾儘僶僋僠儞丄儕僝僶僋僠儞丄僔僝僉僯儞乯側偳傕棙梡偡傞偙偲偑壜擻偱偁傞丅
偝傜偵丄僇僥僐乕儖傾儈儞恄宱揱払暔幙傗怉暔桼棃偺儌僲僇僥僐乕儖偵壛偊丄僋僄儞巁傪僔僨儘僼僅傾偲偟偰棙梡偡傞偙偲傕壜擻偱偁傞丅
傑偨丄 P. aeruginosa僎僲儉偵偼丄3偮偺僿儉庢傝崬傒宱楬乮僿儉偼揝尮乯偲1偮偺揝妉摼宱楬偑僐乕僪偝傟偰偄傞丅
杮憤愢偼丄 P. aeruginosa偑梡偄傞偡傋偰偺揝偍傛傃僿儉妉摼宱楬偵娭傢傞暘巕婡峔偵娭偡傞尰嵼偺抦尒傪梫栺偡傞
偙偙偱偼彫惗偑壓恾懡偔偺偦傟偧傟偺僔僨儘僼僅傾偺庢傝崬傒婡峔偺偆偪丄怉暔偵娭學偡傞偲巚傢傟傞僋僄儞巁揝偲僿儉揝偺媧廂婡峔偵偮偄偰偺傒徯夘偡傞丅
乮恾1偺愢柧乯丗僋僄儞巁埶懚偺揝暘庢傝崬傒宱楬
椢擽嬠偼僋僄儞巁傪僄僉僜僔僪僼僅傾偲偟偰棙梡偡傞偙偲偑偱偒傞(Marshall et al.丆2009乯丅
僋僄儞巁戞擇揝暋崌懱偼丄TBDT偱偁傞FecA乮PA4825乯偵傛偭偰擣幆偝傟丄儁儕僾儔僗儉偵桝憲偝傟傞丅
僋僄儞巁戞擇揝暋崌懱偼丄梟塼拞偺暯峵忬懺偱嫟懚偡傞丄壔妛検榑揑偵條乆側暋崌懱偺偐側傝戝偒側僇僥僑儕乕傪昞偟偰偄傞丅
偙傟傜偺嶖懱偼丄嬥懏偲偺寢崌偵悈暘巕偑娭梌偟偰偄傞偙偲傕偁傝丄廳崌偡傞偙偲傕壜擻偱偁傞丅
廬偭偰丄TBDT FecA偑擣幆偡傞戞擇揝-僋僄儞巁暋崌懱偺壔妛検榑傪寛掕偡傞偙偲偼崲擄偱偁傞偑丄暥專揑偵偼戞擇揝-僕僋儗乕僩宆偑擣幆偝傟傞偙偲偑摉慠偲偝傟偰偄傞傛偆偱偁傞丅
fecA偺揮幨偼丄僔僌儅儁傾ECF/傾儞僠僔僌儅FecI/FecR偵傛偭偰惂屼偝傟偰偄傞丅
儁儕僾儔僘儉偵擖傞偲丄儁儕僾儔僘儉偺僼僃儗僪僉僔儞PcoA偵傛傞揝偺娨尦傪敽偆儊僇僯僘儉偱丄揝-僋僄儞巁暋崌懱偑夝棧偡傞偲峫偊傜傟偰偄傞丅
戞堦揝偺傒偑撪枌偺FeoAB傪宱桼偟偰撪枌傪堏摦偡傞丅
乮恾2偺愢柧乯丗僿儉偺愛庢
尰嵼丄椢擽嬠偱偼丄Phu乮Pseudomonas heme uptake乯丄Has乮heme-acquisition system乯丄PA1300-1302宯乮Smith & Wilks, 2015乯偲偄偆3偮偺僿儉妉摼宯偑婰嵹偝傟偰偄傞丅
PA1302偼丄HxuC偲偟偰傕抦傜傟傞TBDT傪僐乕僪偡傞偲梊應偝傟傞堚揱巕偱丄嵟嬤HxuA偲夵柤偝傟丄偙偺僩儔儞僗億乕僞乕傪庢傝姫偔崿棎偵攺幵傪偐偗偰偄傞丅
Phu偍傛傃Has宯偼旕忢偵傛偔摿挜晅偗傜傟偰偄傞偺偵懳偟丄僿儉TBDT撪偺PA1302偺暘椶偼傑偩晄柧偱偁傞丅
Hxu偼丄梀棧僿儉偺桝擖偵娭梌偡傞Haemophilus influenzae偺HxuC僩儔儞僗億乕僞乕偲偺嫮偄憡摨惈偵婎偯偄偰丄僿儉TBDT偲偟偰嵟弶偵摿掕偝傟偨丅
嵟嬤偺尋媶偵傛傞偲丄HxuA偼崅擹搙乮20兪M乯偺僿儉懚嵼壓偱夁忚惗嶻偝傟丄HxuA偺揮幨偼僔僌儅乛傾儞僠僔僌儅場巕懳傪僐乕僪偡傞HxuI偲HxuR偺惂屼壓偵偁傞偲悇應偝傟偰偄傞丅
椢擽嬠偺僿儉儂儊僆僗僞僔僗偵偍偗傞HxuA偺栶妱偵娭偡傞僨乕僞偼丄戙傢傝偵丄桝憲偺栶妱傛傝傕傓偟傠僔僌僫儕儞僌偺栶妱傪帵偡孹岦偵偁傞丅
Phu僔僗僥儉偼丄梀棧僿儉傪嵶嬠儁儕僾儔僗儉偵桝擖偡傞偙偲偑偱偒傞TBDT PhuR偲丄僆儁儘儞phuSTUVW偵傛偭偰僐乕僪偝傟傞僞儞僷僋幙偱峔惉偝傟偰偄傞丅
phuR偼揝暘晄懌偺忦審壓偱夁忚敪尰偡傞偑丄偦偺揮幨惂屼宯偼傑偩摿掕偝傟偰偄側偄丅
僿儉偼PhuR偵傛偭偰儁儕僾儔僗儉偵庢傝崬傑傟傞偲丄儁儕僾儔僗儉寢崌僞儞僷僋幙PhuT偵寢崌偟丄撪枌ABC僩儔儞僗億乕僞乕PhuUV偵塣偽傟傞丅
嵶朎幙偵擖傞偲丄僿儉偼PhuS僞儞僷僋幙偵寢崌偡傞丅HemO偼PhuS-僿儉暋崌懱偲憡屳嶌梡偟丄奜場惈偺僿儉傪價儕儀儖僕儞IXß偲價儕儀儖僕儞IX兟偵暘夝偡傞丅
嵶朎撪偵ß偍傛傃兟價儕儀儖僕儞偑懚嵼偟側偄偲丄Has宯偺僿儌僼僅傾HasAp偺嵶朎奜擹搙偑掅壓偡傞偙偲偐傜丄ß偍傛傃兟價儕儀儖僕儞偺惂屼揑栶妱偑帵嵈偝傟傞丅
phuW偑敪尰偡傞僞儞僷僋幙偺栶妱傗嬊嵼偼尰嵼偺偲偙傠晄柧偱偁傞偑丄偙偺堚揱巕偼僿儉妉摼偵娭梌偡傞儕億僞儞僷僋幙僼傽儈儕乕偺壖愢偱偁傞ChaN條僞儞僷僋幙傪僐乕僪偡傞傕偺偲梊應偝傟偰偄傞丅
Has僔僗僥儉偼丄HasAp僿儌僼僅傾偲偄偆丄娐嫬偐傜僿儉傪曔懆偟丄TBDT HasR偵傛偭偰擣幆偝傟傞彫偝側暘斿僞儞僷僋幙傪娷傓偲偄偆揰偱旕忢偵捒偟偄丅
奜枌儗儀儖偱偺偙偺憡屳嶌梡偵傛傝丄僿儉偑儁儕僾儔僘儉偵堏摦偟丄偦偙偱PhuT偵寢崌偟偰Phu宱楬偵壛傢傞丅
Phu宯偲偼堎側傝丄Has宯偼僔僌儅/傾儞僠僔僌儅場巕HasI/HasS偵傛偭偰惂屼偝傟偰偄傞丅HasAp偺嵶朎奜娐嫬傊偺暘斿偼丄HasEDOpmM攔弌宯偵埶懚偟偰偄傞丅
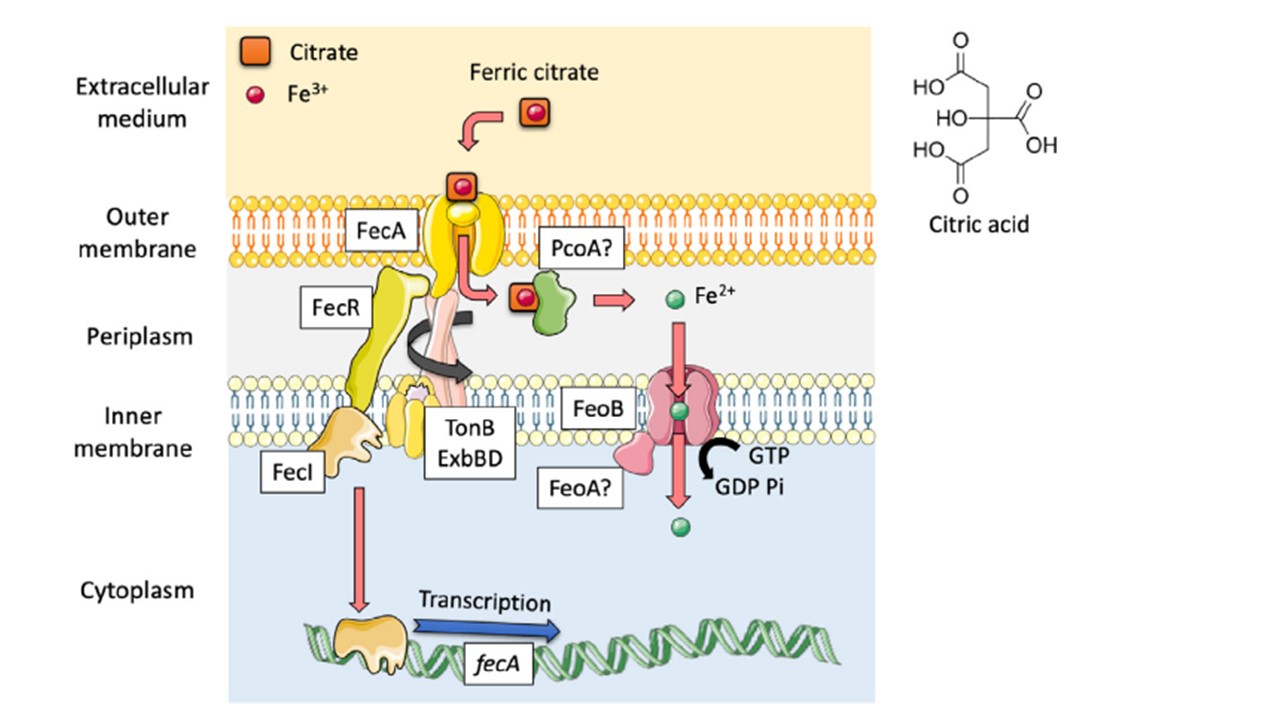
恾1丗乽僋僄儞巁揝乿媧廂儌僨儖
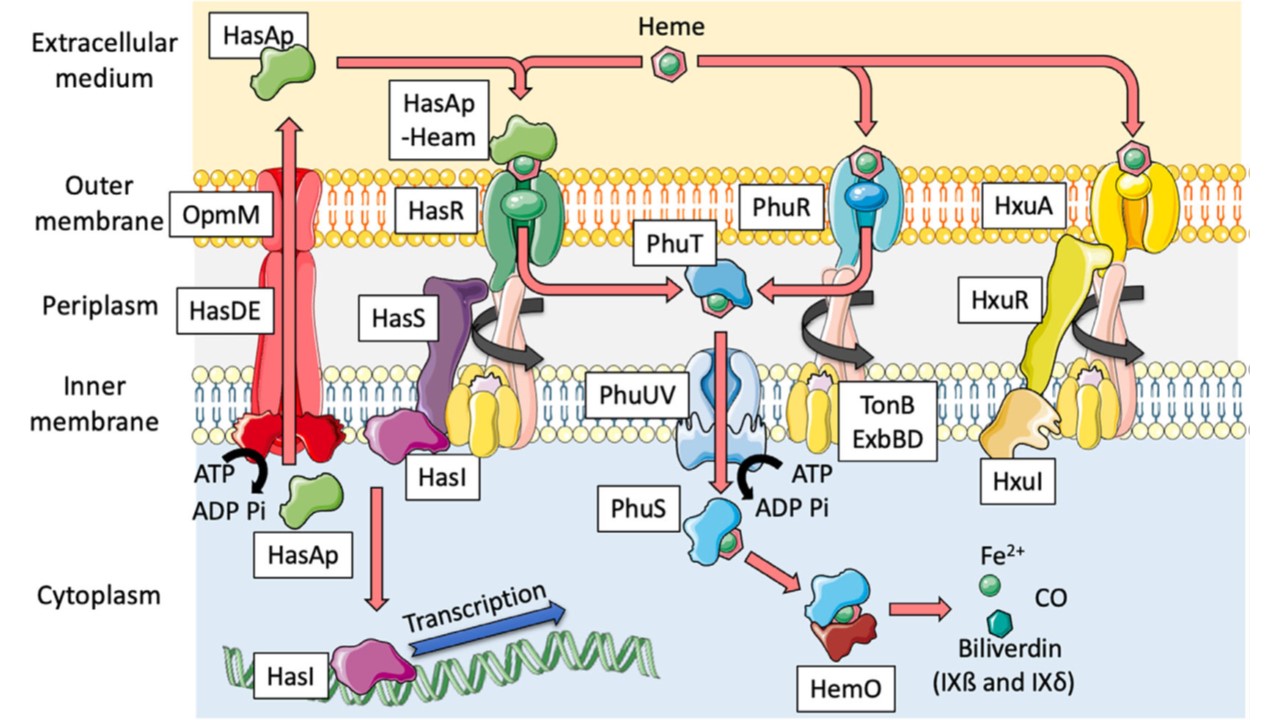
恾2丗乽僿儉揝乿媧廂儌僨儖