イネいもち病菌Magnaporthe oryzaeに対する免疫応答の増強による抵抗性の誘導について
雑誌 「Rice」 に掲載されたこの論文はいささか衝撃的である。イモチ菌に感染したイネの葉が、免疫応答して、感染細胞周辺が細胞死して感染斑を形成して、そこで感染の拡大を食い止める、ということはこれまでもよく知られていた。
この論文では、この感染斑が鉄による活性酸素の放出によるフェロトーシス(feroptosis)によるものであるとしている。
これまで日本のイネの研究者は、水田では二価鉄が十分にあるのでイネは鉄欠乏になるはずがないと考えてきた。しかし,イネの発育段階で、低温寡照や窒素過多などによって、イネが徒長する条件下では、イネの体内では潜在的な鉄欠乏が起こっている可能性がある。
そういう場合はイモチ菌に感染すると鉄によるフェントン反応による活性酸素の発生が間に合わないために、急速な感染糸の細胞間拡散が行われるのであろう。
私見では、この研究は植物栄養学と植物病理学の接点を切開した良質の研究であると思う。
技術的には低温寡照が続くときには水稲には鉄剤を予防的に葉面散布することが勧められることになる。
イモチ病菌(Magnaporthe oryzae)に対する免疫応答の増強による抵抗性の誘導について
Sanchez‑Sanuy et al. Rice (2022) 15:68
https://doi.org/10.1186/s12284-022-00609-w
(概要)
鉄は、植物の成長と発達に必要な必須栄養素である。
また、鉄の利用可能性は、植物の病害抵抗性にも影響を与える可能性がある。
しかし、鉄の利用可能性と免疫に対する植物の反応に関わる分子機構は、それぞれ別々に研究されてきた。
本研究では、イネを高鉄分に曝露することで、いもち病の原因菌であるMagnaporthe oryzaeの感染に対する抵抗性が増強されることを見いだした。
RNA-Seq 解析により、鉄処理イネのいもち病抵抗性は、病原体感染時の防御関連遺伝子(Pathogenesis-Related genes)のスーパーインダクションと関連していることが明らかとなった。
ジテルペン系ファイトアレキシンとフラボノイド系ファイトアレキシンサクラネチンの生合成に関わる遺伝子の発現量を調べたところ、M. oryzae感染時に鉄処理植物においてファイトアレキシンのレベルが増加することとよく相関していた。
病原体感染時には、鉄処理植物で過酸化脂質が無処理植物に比べて高くなることもわかった。
さらに、M. oryzae 感染が鉄の恒常性維持に重要な役割を果たす遺伝子の発現を調節していることも明らかにした。
また、M. oryzae感染葉の組織化学的解析から、鉄に曝されたイネでは、菌の侵入部位(被膜など)の近傍の細胞で鉄と活性酸素の共局在が確認された。
これらの結果は、鉄処理したイネが病原性M. oryzaeの感染に対してフェロプートシスが関与していることを支持するものである。
イネにおける鉄シグナルと免疫シグナルの相互関係を理解することは、イネのいもち病抵抗性を向上させるための新たな戦略の開発につながることが期待される。
以下これまでの研究紹介(部分訳)
根に比べ、葉の組織における鉄の輸送と恒常性の制御機構に関する知見はまだ限られている。
鉄の栄養状態は、植物の病害抵抗性に影響を与える可能性がある(Herlihy et al.2020、Liu et al.2007)。
葉面病原体がこの重要な元素を獲得するために宿主組織に全面的に依存していることを考慮すると、病原体感染時には、宿主と病原体の間で鉄の獲得競争が起こっていることになる。
植物と病原体の相互作用において、植物は病原体の毒性を弱めるために様々な戦略をとるかもしれない。
一方、植物は鉄を病原体から隔離することで、鉄を保有する戦略をとるかもしれない。この現象は、もともと動物で報告され、「栄養免疫」と呼ばれている(Weinberg 1975)。
一方、宿主植物は、侵入した病原体にとって有害な鉄を局所的に蓄積することによって、鉄の毒性を利用するかもしれない(Herlihy et al.2020)。
また、植物病原菌は、高親和性鉄結合性シデロフォアの分泌など、宿主植物から鉄を獲得する戦略を展開する可能性もある(Herlihy et al.)
宿主や病原体がこのような戦略をとるかどうかが、相互作用の結果を左右すると思われる。
鉄の枯渇を感知することは、植物が病原菌の脅威を認識し、免疫反応を活性化させるメカニズムとして提唱されている(Herlihy et al.)
この考え方を支持するものとして、鉄欠乏させたシロイヌナズナでは、Dickeya dadantii や Botrytis cinerea の感染に対する抵抗性が観察された (Kieu et al. 2012)。
他の研究では、鉄欠乏トウモロコシはColletotrichum graminicola感染時に活性酸素を生成できず、この菌類病原体に対する感受性の増加と相関することが見出された(Ye et al.)。
一方、Curvularia lunataによる葉面斑点病の感染時にトウモロコシで報告されたように、局所的な過剰鉄が活性酸素バーストを活性化し、病原菌に抵抗することが知られている(Fu et al.)。
現在のところ、植物における鉄のホメオスタシスと免疫反応を調整するクロストーク機構は、特に作物種において十分に理解されていない。
イネ(Oryza sativa L.)は世界で最も重要な作物の一つであり、その生産はMagnaporthe oryzaeという真菌によって深刻な影響を受けている。この菌は、イネいもち病の原因菌である(Wilson and Talbot 2009; Fernandez and Orth 2018)。
これまでの研究において、Fe3+と活性酸素(e.g. H2O2)がM. oryzaeの非毒性株による感染時にイネの葉に蓄積することが報告されている(Dangol et al.2019)。
本研究では、高濃度鉄処理がいもち病抵抗性に及ぼす影響について、分子・細胞レベルで検討した。
RNA-Seq解析により、イネを高濃度鉄に短時間曝露すると、病原菌関連(PR)遺伝子の発現を含む防御応答がより強く誘導されることが明らかとなった。
また、鉄処理イネでは、M. oryzae 感染によりフィトアレキシンの生合成遺伝子が強く誘導され、このことは、フィトアレキシンの高蓄積とよく相関していた。
このことは、鉄処理イネにおけるフィトアレキシンの高蓄積とよく相関していた。
鉄処理イネでは,防御遺伝子の活性化がM. oryzae感染に対する抵抗性に関連していることが示唆された。
また,病原菌の感染は,イネの葉の鉄のホメオスタシスに関与する遺伝子の発現を調節し,鉄含量(全 鉄,葉上鉄)を変化させることが明らかとなった。
鉄処理イネに強毒性 M. oryzae を感染させると、組織化学的染色により活性酸素と鉄が共局在化し、M. oryzae 感染時に脂質過酸化レベルが上昇することが明らかとなった。
フェロトシス阻害剤であるフェロスタチン-1(Fer-1)投与により鉄の蓄積が抑制されたことから、鉄処理イネではフェロトシスがいもち病抵抗性に関与していることが示唆された。
これらの結果は、イネにおける鉄のシグナル伝達と免疫シグナルの関係をさらに裏付けるとともに、イネのいもち病抵抗性付与におけるフェロトシスの重要性を浮き彫りにするものである。
以下図の説明(紙面の制限のために、9枚ある図のうち、図6,図8,図9のみを訳した)
図6 高濃度鉄供給下で生育したイネの葉におけるフィトアレキシンの蓄積。
A イネのメチルエリスリトールリン酸(MEP)およびジテルペノイド系フィトアレキシンの生合成経路。
M. oryzae感染により、対照植物および高Fe供給植物で発現が上昇する遺伝子を紫色で示した。
これらの遺伝子の正式名称と発現の詳細は、Additional file 6: Table S5 に記載されている。
B コントロール植物とHigh-Fe植物、モック接種植物とM. oryzae接種植物の葉における発現量(行スケールのFPKM、100万リードあたりの断片数)を示すヒートマップ(48hpi)。
遺伝子発現は、淡黄色(発現が少ない)から茶色(発現が多い)へと表現されている。
C M. oryzae胞子を接種したControlとHigh-Fe植物の48hpiと72hpiの葉におけるジテルペノイド系フィトアレキシンとサクラネチンの蓄積:モミラクトン(AおよびB)、フィトカッサン(B、C、E)、フラボノイド系フィトアレクシンサクラネチン(-、モック接種;+、M. oryzae接種)。
G3P、グリセルアルデヒド-3-リン酸;DXP、1-デオキシ-D-キシルロース5-リン酸;MEP、2-C-メチル-D-エリスリトール4-リン酸;CDP-ME、4-(シチジン5´-ジホスホ)-2-C-メチル-D-エリスリトール;CDP-ME2P、2-ホスホ-4-(シチジン5´-ジホスホ)-2-C-メチル-D-エリスリトール。MECDP, 2-C-methyl-D-erythritol 2,4-cyclodiphosphate; HMBDP, 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate; IPP, isopentenyl diphosphate; DMAPP, dimethylallyl diphosphate; GGDP, geranylgeranyl diphosphate; CDP, copalyl diphosphateの4種が挙げられる。
図 8 対照イネおよび高 Feの葉における活性酸素と Fe の蓄積。
A 蛍光プローブ H2DCFDA を用いて葉中の活性酸素(H2O2)を検出した。
モックを接種した植物(mock)とM. oryzaeを接種した植物(48 hpi)の葉(最も若く展開した葉)を調べた。棒は2 mmに相当する。
右図、Image J ソフトウェアを用いた活性酸素蛍光の定量化(3回の独立実験、100フィールド、各)。
B 模擬接種した Control-Fe 植物(上段)と M. oryzae に感染させた High-Fe 植物(下段)の葉における鉄検出(48 hpi)。
最も若い葉(第3葉)をDAB染色(茶色)した後、Perls染色(青色)を行った。棒グラフは50μmに対応する。St, 気孔, Ap: 付着葉。
図 9 M. oryzae 胞子を接種した Low-Fe, Control-Fe, High-Fe イネの葉における鉄検出(48 hpi)。
葉はフェロプ トシス阻害剤フェロスタチン(Fer-1)で処理した(+ Fer-1)、または処理しない(- Fer-1)。
イネの葉(最も若い展開葉)をPerls染色して、Feを検出した(青色)。
表示された画像は、2つの独立した実験の代表である。棒グラフは100μmに相当する。
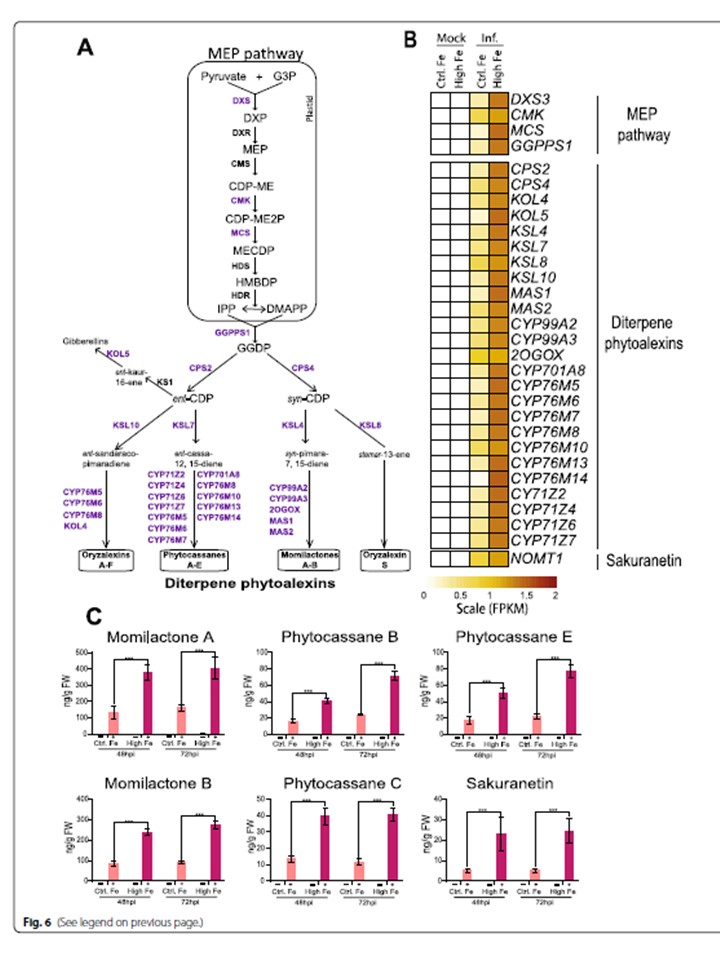
図6.
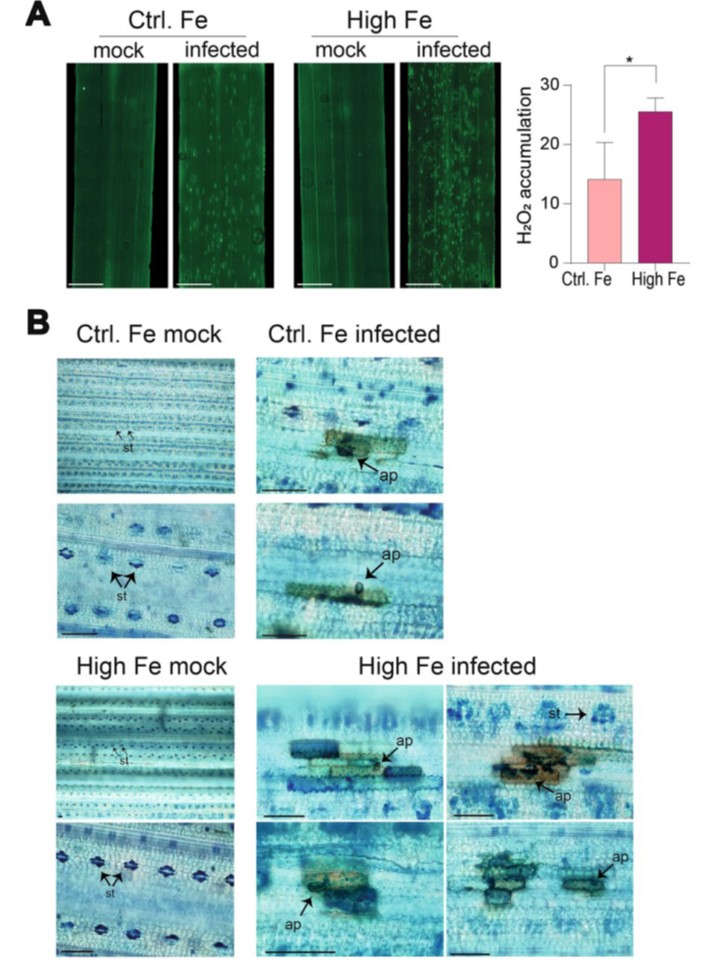
図8.
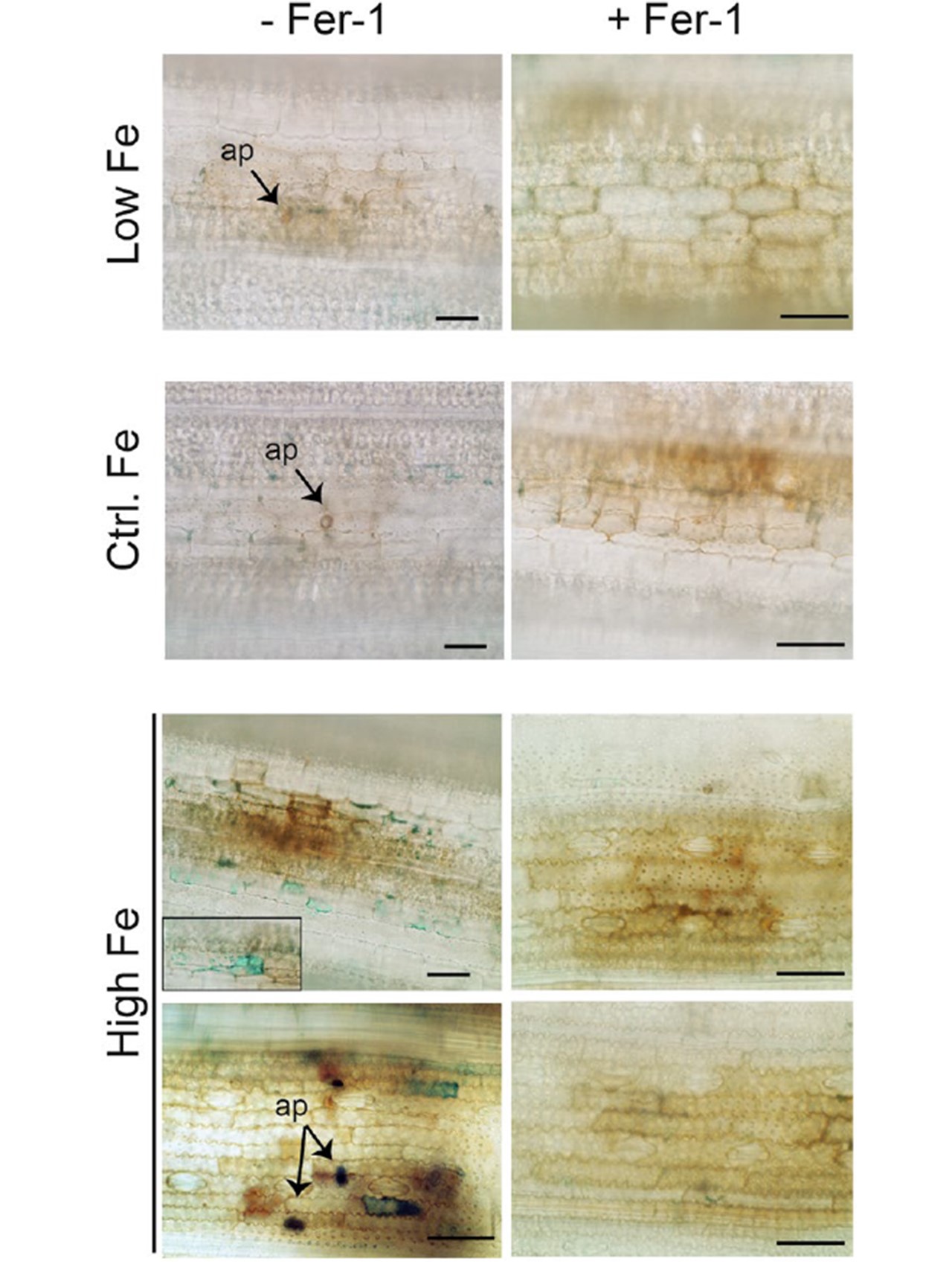
図9.