怉暔偼偳偺傛偆偵嵶朎撪偺揝偺儂儊僆僗僞僔僗傪嵞挷惍偟偰偄傞偺偐
埲壓偺儈僯儗價儏乕偼側偐側偐傛偔彂偗偰偄傞偺偱丄慡暥栿偟偨丅
俀侽侾俋亅俀侽俀侾擭娫偵弌斉偝傟偨丄怉暔偺揝塰梴娭學偺廳梫榑暥傪傛偔栐梾偟偰丄傑偲傔偰偄傞偲巚偆丅
儈僯儗價儏乕丗
怉暔偼偳偺傛偆偵嵶朎撪偺揝偺儂儊僆僗僞僔僗傪嵞挷惍偟偰偄傞偺偐
丂丂
How Plants Recalibrate Cellular Iron Homeostasis
丂丂
僀僓儀儖丒僋儕僗僥傿乕僫丒儀儖儉僨僗丄僂僅儖僼僈儞僌丒僔儏儈僢僩
丂丂
Plant Cell Physiol 63(2): 154-162 (2022) https://academic.oup.com/pcp
丂丂
丂丂
亙傑偊偑偒亜
丂丂
揝偺嫙媼晄懌偼怉暔偵怺崗側惂栺傪梌偊丄揝偺棙梡壜擻惈偑掅偄惗懅抧偐傜揝偺庢傝崬傒婡峔偑旕岠棪側庬偑攔彍偝傟傞偺偱丄擾嬈惗懺宯偵偍偗傞廂検懝幐傪堷偒婲偙偡丅
傑偨丄揝偺寚朢偼恖娫偺寬峃偵傕怺崗側嫼埿傪梌偊偰偄傞丅
揝偺愛庢晄懌偵傛傞昻寣偼丄悽奅偺4恖偵1恖偑滊姵偟偰偄傞偲尵傢傟偰偄傞丅
廬偭偰丄怉暔偑搚忞偲嵶朎偺娫偺戝偒側岡攝偵媡傜偭偰揝傪妉摼偡傞儊僇僯僘儉傗丄怉暔懱撪偱揝偑偳偺傛偆偵暘晍偟偰偄傞偐傪棟夝偟丄怘梡怉暔懱撪偺揝擹搙傪崅傔傞愴棯傪棫偰傞偙偲偑晄壜寚偱偁傞丅
怉暔偑嵶朎撪偺揝偺儂儊僆僗僞僔僗傪挷惍偡傞僾儘僙僗傪尋媶偟偨寢壥丄僔僌僫儖揱払僲乕僪偲夞楬偑嬃偔傎偳暋嶨側僷僘儖偺傛偆偵棈傒崌偭偰偄傞偙偲偑柧傜偐偵側偭偨丅偙偺夞楬偼丄昦尨嬠傗岝丄塰梴偺桳柍丄pH側偳偺晽搚揑梫場側偳丄懠偺娐嫬偺庤偑偐傝傪抦妎偟丄揱払偡傞偙偲偲棈傒崌偭偰偄傞偺偩丅
杮帍(Plant Cell Physiol)偺嵟嬤偺Spotlight崋偱偼丄怉暔揝尋媶偺嵟愭抂傪傑偲傔偨憤愢偑宖嵹偝傟丄偙偺暘栰偱嵟傕妶敪偱丄媍榑偺梋抧偑偁傞嵟傕廳梫側僩僺僢僋偑栐梾偝傟偰偄傞丅
偙偙偱偼丄偙偺憤愢廤偺弌斉擔埲崀偵曬崘偝傟偨夋婜揑側尋媶傪庢傝忋偘丄怉暔偺揝塰梴偵娭偡傞変乆偺棟夝傪曄偊傞傛偆側丄巋寖揑偱塭嬁椡偺偁傝偦偆側尋媶偵徟揰傪摉偰傞丅
丂丂
丂丂
亙揔嵽揔強丄揔帪丄揔検丗揝暘曗媼偺嬯楯榖亜
丂丂
揝偼壙悢偑曄壔偟傗偡偄偨傔丄岝崌惉傗屇媧偺揹巕揱払宯偺廳梫側峔惉梫慺偱偁傝丄惗柦堐帩偺偨傔偺柍悢偺僾儘僙僗偵偍偄偰晄壜寚側曗場巕偱偁傞丅怉暔偑揝傪棙梡偱偒傞偐偳偆偐偼搚忞偺pH偲巁慺擹搙偵戝偒偔埶懚偟丄傎偲傫偳偺応崌丄偦偺嫙媼検偼丄嵟揔側惗堢偺偨傔偺昁梫検傪壓夞偭偰偄傞丅
帺慠奅偱偼偙偺帠偑乽庬乿偺暘晍傪惂尷偟丄懡偔偺擾嬈娐嫬偱偼嶌暔偺惗堢傪惂尷偟偰偄傞丅
揝偺嫙媼偑晄廫暘側応崌丄梩偺僋儘儘僔僗丄惉挿偺慾奞丄懱椡偺掅壓側偳偑婲偙傞丅
傑偨丄揝偑夁忚偵側傞偲丄椺偊偽扻悈忦審壓偱偼妶惈巁慺偺惗惉偑憹偊偰巁壔揑側忈奞偺惗惉偵偮側偑傞丅
揝埶懚惈僞儞僷僋幙偺岆嶌摦偲妶惈巁慺偺晄嬒峵偺椉曽傪夞旔偡傞偨傔偵丄嵶朎偺揝儗儀儖傪揔愗偵挷惍偡傞偙偲偼丄乽偪傚偆偳偄偄忦審傪乿偲偄偆崲擄側儈僢僔儑儞偱偁傞丅
偙偺偨傔丄搚忞拞偺揝偺棙梡壜擻検偑掅偡偓傞丄偁傞偄偼崅偡偓傞応崌丄偦傟傪曗偆偨傔偵丄揝偺庢傝崬傒偲暘攝傪挷惍偡傞偙偲傪栚揑偲偟偨堦楢偺斀墳偑怉暔偵偼昁梫偲側傞丅
揝偺庢傝崬傒偼丄揝偺媧廂偲儂儊僆僗僞僔僗傪拠夘偡傞僞儞僷僋幙偺検傪挷惍偡傞僩儔儞僗嶌梡場巕丄惂屼儁僾僠僪丄儐價僉僠儞儕僈乕僛側偳偺旕忢偵暋嶨側僱僢僩儚乕僋偵傛偭偰惂屼偝傟偰偄傞丅
僔儘僀僰僫僘僫偱偼丄偙偺惂屼僱僢僩儚乕僋偺僉乕僾儗僀儎乕偼丄揝僙儞僒乕偲悇掕偝傟傞BRUTUS乮BTS乯偺敪尰傪惂屼偡傞clade IVc bHLH僞儞僷僋幙IAA-LEUCINE RESISTANT 3乮ILR3乯/bHLH105丂偍傛傃丂bHLH115偲丄IRONMAN/PE-UPTAKE-IDUCING PEPTIDE (IMA/FEP) 偲屇偽傟傞惂屼儁僾僠僪僼傽儈儕乕乮Gao et al.2020,Grillet et al.2018,Hirayama et al.2018) 偱偁傞丅
暿偺bHLH僞儞僷僋幙丄UPSTREAM REGULATOR
IRT1 (URI/bHLH121)偼丄ILR3/bHLH105丄IRON DEFICIENCY TOLERANT 1 (IDT1)/bHLH34丄bHLH104偍傛傃bHLH115偲憡屳嶌梡偟偰丄clade Ib bHLH僞儞僷僋幙偺揮幨傪惂屼偟偰偄傞丅
暿偺bHLH僞儞僷僋幙丄UPSTREAM REGULATOR OF IRT1乮URI/bHLH121乯偼丄ILR3/bHLH105丄IRON DEFICIENCY TOLERANT 1乮IDT1乯/bHLH34丄bHLH104丄bHLH115偲憡屳嶌梡偟偰丄clade Ib bHLH僞儞僷僋幙偍傛傃偺揮幨傪惂屼偟偰偄傞丅捈愙傑偨偼娫愙揑偵丄儅僗僞乕儗僊儏儗乕僞乕FIT/bHLH29偼丄僋儗乕僪Ib僞儞僷僋幙偲僿僥儘僟僀儅乕傪宍惉偟偰丄崻偵偍偗傞揝庢傝崬傒堚揱巕偺僒僽僙僢僩偺揮幨傪桿摫偡傞乮Gao et al. 2020, Kim et al. 2019, Lei et al. 2020)丅
嵟廔揑偵丄偙偺僇僗働乕僪偼丄僆僉僔悈巁壔暔偺僾儘僩儞壔丄僉儗乕僩壔丄娨尦偵傛偭偰丄揝偺棙梡棪傪崅傔傞堦楢偺僾儘僙僗傪堷偒婲偙偡丅
偙傟偑丄愴棯- I 偲屇偽傟傞揝偺庢傝崬傒奣擮偱偁傞 (R丯omheld and Marschner 1986)丅
偙傟偵懳偟丄僀僱壢怉暔偼丄娨尦揑側揝偺庢傝崬傒丄偡側傢偪愴棯- I
曽幃偐傜堩扙偟偰丄揝偵崅搙偵恊榓惈偺僼傿僩僔僨儘僼僅傾偵寢崌偟偨戞擇揝偺宍偱庢傝崬傓僔僗僥儉傪嵦梡偟偰偄傞丅偙傟傪愴棯-II偲屇傫偱偄傞丅
廬棃丄偙偺2偮偺僔僗僥儉偼屳偄偵憡梕傟側偄偲峫偊傜傟偰偄偨丅偟偐偟
嵟嬤丄偙偺2偮偺愴棯偺嬫暿偼庛傑傝偮偮偁傞乮壓婰嶲徠乯丅
(偟偐偟丄僀僱壢怉暔偵偼桿摫惈揝娨尦峺慺妶惈偑側偄側偳偺庡側堘偄偑偁傝丄宯摑揑偵暘棧偟偨2偮偺堎側傞揝偺庢傝崬傒愴棯偲偄偆僐儞僙僾僩偼埶慠偲偟偰桳岠偱偁傞乮Kobayashi 2019乯丅
偙傟傑偱偺杮帍偵傛傞曪妵揑側儗價儏乕偺僼僅儘乕傾僢僾偲偟偰丄変乆偼
偙傟傜偺暘栰偱丄摿廤崋敪峴屻偺2擭娫偵曬崘偝傟偨丄摿偵廳梫側抦尒傪徯夘偡傞丅
偙偺暘栰偺暥專傪曪妵揑偵挷嵏偡傞偲偄偆傛傝偼丄傓偟傠丄揝偺妉摼偲儂儊僆僗僞僔僗偺奣擮偺夝庍偺巇曽傪戝偒偔曄偊傞偙偲偵側偭偨堦楢偺尋媶偵摿偵拲栚偟偨偄丅
偙偺儗價儏乕偺斖埻奜偱偼偁傞偑丄偙偙偱偼徻偟偔愢柧偟偒傟側偐偭偨偑丄堄媊偺崅偄尋媶傪埲壓偵偄偔偮偐徯夘偟偨偄丅
揝寚朢斀墳偺惂屼偵偍偗傞OsACO1偺moonlighting Function乮寧岝梺婡擻丠乯(Senoura et al. 2020)丄揝妉摼婡峔偺嵶朎撪僟僀僫儈僋僗(Martín-Barrn, 1996)丄崌惉僼傿僩僔僨儘僼僅傾偵傛傞揝偺巤旍(Suzuki et al. 2021)偍傛傃庬巕傊偺揝偺堏峴(Ch. et al. 2021)丅
丂丂
丂丂
亙OsFIT偲OsIMA偼僔儘僀僰僫僘僫偺婡擻揑儂儌儘僌偱偁傞亜
丂丂
揝寚朢偵懳偡傞揟宆揑側斀墳偑偡傋偰徚幐偟偨僩儅僩曄堎懱T3238fer偺廳搙偺僋儘儘僔僗昞尰宆偼丄揝寚朢偺惂屼偺婎斦偲側傞暘巕婡峔偺夝柧偺巒傑傝偲側偭偨丅
偙偺曄堎懱偼1970擭戙弶摢偵弶傔偰曬崘偝傟丄偦偺屻丄愼怓懱6忋偺崻偵嬊嵼偡傞場巕偵楎惈曄堎偑偁傞偙偲偑傢偐偭偨(Brown et al. 1971, Ling et al. 1996)丅
FER偺僋儘乕僯儞僌偵傛傝丄曄堎偼揮幨場巕SlbHLH85偲娭楢晅偗傜傟偨(Ling et al. 2002)丅僔儘僀僰僫僘僫偺僆儖僜儘僌偱偁傞fer, fer-like iron deficiency-induced transcription FACTOR (FIT/bHLHLH85) 偺僔儘僀僰僫僘僫偺僆儖僜儘僌偱偁傞FACTOR (FIT/bHLH29)偑丄傢偢偐2擭屻偵摨掕偝傟偨(Jakoby丂et al. 2004, Colangelo and Guerinot 2004, Yuan et al. 2005)丅偦傟埲棃丄峀斖埻偵摿惈昡壙偝傟偰偄傞乮Lingam et al.2011,
Wu and Ling 2019, Gratz et al. 2019, Schwarz and Bauer 2020)丅
FIT偼4偮偺僋儗乕僪Ib bHLH僞儞僷僋幙乮bHLH38丄bHLH39丄bHLH100丄bHLH101乯偺偆偪1偮偲僿僥儘2検懱傪宍惉偟丄廳梫側堚揱巕傪惂屼偟偰偄傞丅
偦偺拞偵偼丄愴棯-I偺庡梫側揝媧廂僾儘僙僗傪攠夘偡傞堚揱巕傕娷傑傟偰偄傞丅偨偲偊偽丄巁惈壔(AHA2)丄僉儗乕僩壔 (F6亴H1, S8H, CYP82C4)丄娨尦 (FRO2)丄揝偺桝憲 (IRT1) 側偳偺堚揱巕偱偁傞丅乮Yuan et al.2008丄Wang et al.2013乯丅
偙傟傑偱丄僀僱偺傛偆側愴棯-II宆怉暔偲僩儅僩傗僔儘僀僰僫僘僫偺傛偆側愴棯-I宆怉暔偲偱偼丄惂屼僇僗働乕僪偑戝偒偔堎側傞偲峫偊傜傟偰偄偨丅偟偐偟
揝偺儂儊僆僗僞僔僗偺惂屼偼丄偙偺2偮偺儌僨儖怉暔娫偱偼丄寢嬊偺偲偙傠丄偦傟傎偳堘偄偼側偄傛偆偩乮Grillet and Schmidt 2019乯丅
偙偺尒夝偼丄嵟嬤偵側偭偰傛傝懡偔偺巟帩傪摼偨丅2偮偺僌儖乕僾偑撈棫偟偰
FIT偺婡擻揑側僀僱儂儌儘僌偱偁傞OsbHLH156傪摨掕偟偨乮Liang et al.2020丄Wang et al.2020乯丅
AtFIT傗偦偺僩儅僩偺儂儌儘僌SlFER偲摨條偵丄OsFIT偼clade IIIa bHLH僞儞僷僋幙偱偁傝丄僋儗乕僪Ib僞儞僷僋幙偱偁傞IRO2 (bHLH56; Ogo et al., 2006, Liang et al. 2020, Wang et al. 2020)偲僿僥儘僟僀儅乕傪宍惉偟偰偄傞丅
OsFIT偵曄堎偑偁傞偲丄怉暔偺僼傿僩僔僨儘僼僅傾偺崌惉偲桝憲偵娭傢傞偄偔偮偐偺廳梫側堚揱巕偺敪尰偑掅壓偡傞偙偲偐傜丄OsFIT偑愴棯-II宆揝媧廂偺桿摫偵廳梫偱偁傞偙偲傪帵偟偰偄傞丅
傑偨丄OsFIT傪攋夡偡傞偲丄僀僱偺揝偺庢傝崬傒僔僗僥儉偺愴棯-I偺峔惉梫慺偱偁傞OsIRT1偺敪尰傕掅壓偟偨丅
OsFIT偺敪尒偵傛傝丄旐巕怉暔偺揝偺儂儊僆僗僞僔僗傪惂屼偡傞僱僢僩儚乕僋偑柧傜偐偵側偭偨丅
傑偨丄僀僱偲僔儘僀僰僫僘僫偑揝偺儂儊僆僗僞僔僗傪惂屼偡傞忋偱旕忢偵傛偔帡偰偄傞偙偲傕柧傜偐偵側偭偨丅偙偺偙偲偼丄揝寚朢墳摎偺惂屼偵偍偗傞僀僱偲僔儘僀僰僫僘僫偺枾愙側椶帡惈傪嫮挷偡傞傕偺偱偁傞丅
儁僾僠僪僼傽儈儕乕IRONMAN (IMA) 偼僔儘僀僰僫僘僫偱嵟弶偵摨掕偝傟丄
嵟嬤丄僀僱偱傕摨條偺栶妱傪壥偨偡偙偲偑帵偝傟偨乮Grillet et al.2018,
Hirayama et al. 2018, Kobayashi et al. 2021)丅柤慜偺桼棃偼 IMA/FEP堚揱巕傪夁忚敪尰偝偣偨堚揱巕慻姺偊怉暔偑揝丒儅儞僈儞拁愊昞尰宆偱偁傞偙偲偵桼棃偡傞乮Grillet et al.2018).
IMA/FEP儁僾僠僪偼丄C枛抂偵儌僠乕僼傪帩偪丄偦傟偼旐巕怉暔慡懱偱崅搙偵曐懚偝傟偰偍傝丄FIT偺忋棳偵嶌梡偡傞偙偲偵傛偭偰丄愴棯-I墳摎傪桿摫偡傞偺偵廫暘偱偁傞乮Grillet et al,丂Kobayashi et al. 2021, Gautam et al. 2021)丅
僔儘僀僰僫僘僫偱偼IMA1/2偺敪尰偼丄bHLH121偲bHLH105/ILR3偵傛偭偰敪尰偑惓偵惂屼偝傟偰偄傞乮Kim et al.2019丄Gao et al.2020乯丅
2偮偺悇掕僀僱儂儌儘僌丄OsIMA1偍傛傃OsIMA2偑丄僔儘僀僰僫僘僫偱堎庬敪尰偝偣傞偲揝寚朢斀墳傪桿摫偡傞偙偲偑帵偝傟偨偑丄僀僱偱偺堚揱巕偺婡擻揑側摿挜晅偗偑寚偗偰偄偨乮Grillet et al.乯 偑偙偺僊儍僢僾偼嵟嬤杽傔傜傟偨丅僔儘僀僰僫僘僫偱娤嶡偝傟偨偺偲摨條偵丄僀僱偺椉儂儌儘僌偺敪尰偼丄僋儗乕僪IVc僞儞僷僋幙乮OsbHLH058偍傛傃OsbHLH059乯偺惂屼壓偱敪尰偑嫮偔桿摫偝傟偨乮Kobayashi傜丄2019乯丅
2偮偺僀僱IMA堚揱巕偺偄偢傟偐傪夁忚敪尰偝偣偨偲偙傠揝偺拁愊偲丄揝庢傝崬傒堚揱巕偺僒僽僙僢僩偺敪尰偑懀恑偝傟偨乮Kobayashi et al.2021乯丅
僔儘僀僰僫僘僫偲僀僱偱偼丄IMA偼bHLH僒僽僌儖乕僾Ib僞儞僷僋幙偲FIT傪僐乕僪偡傞堚揱巕傪惂屼偟偰偍傝丄IMA偼惂屼僇僗働乕僪偺偐側傝忋棳偵埵抲偟偰偄傞乮Grillet et al.2018, Hirayama et al.2018,丂Gautam et al. 2021丄Kobayashi et al. 2021)丅
IMA偑僀僱壢偲旕僀僱壢偺椉曽偱婡擻偟偰偄傞偲偄偆娤嶡偐傜 僀僱壢偺庬偑暘婒偡傞慜偵IMA偑恑壔偟偰偒偨偙偲偑悇應偝傟傞丅
幚嵺丄僀僱壢偺嵟傕屆偄惗暔庬偺僎僲儉偵IMA堚揱巕偑懚嵼偡傞偙偲偼丄
棨忋怉暔偵偍偄偰IMA偑憗偔偐傜弌尰偟偰偄偨偙偲傪帵嵈偟偰偄傞丅
偟偐偟丄僀僱偲僔儘僀僰僫僘僫偱IMA堚揱巕偺悢偑堎側傞棟桼偼晄柧偱偁傞丅
僔儘僀僰僫僘僫僎僲儉偵偼8偮偺IMA堚揱巕偑懚嵼偟丄偦偺偆偪婡擻揑偵夝柧偝傟偰偄傞偺偼3偮乮IMA1,2,3乯偩偗偱偁傞 (Grillet et al. 2018, Hirayama et al. 2018,Gautam et al.2021)丅
僔儘僀僰僫僘僫偱偼丄IMA偑揝偺庢傝崬傒埲奜偺婡擻偱嵦梡偝傟偰偄傞壜擻惈
傗丄摿掕偺IMA儁僾僠僪偼丄揝寚朢斀墳偺摿掕偺峔惉梫慺傪懡彮堎側傞僞乕僎僢僩偱旝挷惍偡傞偨傔偵昁梫偱偁傞壜擻惈偑偁傞丅
嵟傕廳梫側偙偲偼丄OsIMA1傑偨偼OsIMA2偺偄偢傟偐傪夁忚敪尰偝偣傞偲丄
庬巕偺揝暘擹搙傪栺3攞忋徃偝偣丄掅揝暘偵懳偡傞懴惈傪晅梌偡傞偙偲偑偱偒偨偙偲偱偁傞丅
偙傟偼丄揝寚朢惈昻寣偵懳峈偡傞偨傔偵廳梫側丄拞惈傑偨偼傾儖僇儕惈搚忞偱偺惗堢偵桳棙側昞尰宆偱偁傞丅
棨忋怉暔偺揝偺儂儊僆僗僞僔僗偵偍偗傞IMA儁僾僠僪偺廳梫惈偼廫暘偵妋棫偝傟偰偄傞偲巚傢傟傞偑丄IMA偑嵶朎撪偺揝検傪挷愡偡傞暘巕婡峔偼丄偙傟傑偱柧傜偐偵偝傟偰偄側偐偭偨丅
嵟嬤偺尋媶偵傛傝丄bHLH115偍傛傃bHLH105偲嫞崌偟丄C枛抂偺曐懚僪儊僀儞傪夘偟偰BTS偲寢崌偡傞偙偲偱丄IMA偑E3儕僈乕僛BTS傪妘棧偡傞偙偲傪帵偟偨乮Li傜丄2021乯丅
BTS偵傛傞 bHLH115偲bHLH105偺暘夝偺娚榓偼丄IMA傪娷傓揝墳摎惈堚揱巕偺敪尰傪懀恑偡傞偙偲偵傛傝丄bHLH115偲bHLH105偲IMA儁僾僠僪偺娫偱惓偺僼傿乕僪僶僢僋儖乕僾傪宍惉偟偰偄傞乮恾1乯丅
BTS偵傛傞IMA偺暘夝偵傛偭偰丄揝寚朢斀墳偺妶惈壔偲梷惂偺僶儔儞僗偑曵傟丄嵟廔揑偵崻偵傛傞揝偺媧廂偑尭彮偡傞丅
IMA儁僾僠僪偺C枛抂偲丄BTS偲偦偺僀僱偺儂儌儘僌偱偁傞Haemerythrin motif-containing Really Interesting New Gene (RING)- and Zinc-finger protein 1 (HRZ1/HRZ2; Kobayashi et al. 2013乯偲偑椶帡偟偰偄傞偙偲偐傜丄僀僱傗丄偍偦傜偔懠偺 愴棯-II怉暔傕丄摨條偺儊僇僯僘儉偱揝傪姶抦偟 揝儗儀儖偵墳摎偡傞儊僇僯僘儉傪嵦梡偟偰偄傞偲峫偊傜傟傞丅
丂丂丂
丂丂
亙URI偑揝偺僔僌僫儖揱払僷僘儖偺廳梫側僺乕僗傪姰惉偝偣偨亜
丂
AtFIT偲偦偺僆儖僜儘僌偼丄揝偺摑帯懱惂偵懳偟偰揮幨揑偵墳摎偟偰偄傞丅
偙偺偙偲偼丄忋棳偺揮幨場巕偑娭梌偟偰偄傞偙偲傪帵偟偰偄傞丅
僔儘僀僰僫僘僫偱偼丄FIT偺敪尰偼丄4偮偺clade IVc bHLH僞儞僷僋幙乮ILR3/bHLH105, IDT1/bHLH34,丂bHLH104偍傛傃bHLH115乯偵傛傝惂屼偝傟偰偄傞丅偙傟傜偼丄捈愙揑偵clade Ib偺揮幨傪惂屼偟丄娫愙揑偵FIT傪惂屼偡傞乮Gao and Dubos 2021擭憤愢丟Fig.1乯丅
ILR3偼揝偺庢傝崬傒偺妶惈壔場巕偲偟偰傕梷惂場巕偲偟偰傕摥偒丄屻幰偼梷惂揑側揮幨場巕偱偁傞POPEYE偲僿僥儘僟僀儅乕傪宍惉偡傞偙偲偱 揝偺庢傝崬傒傪梷惂偡傞乮PYE; Long et al.2010丄Tissot et al.2019丄Kroh and Pilon 2020乯丅
ILR3傗懠偺bHLH IVc僞儞僷僋幙偑偳偺傛偆偵揝偺庢傝崬傒傪懀恑偡傞偺偐
偼丄嵟嬤傑偱晄柧側傑傑偱偁偭偨丅
儖僔僼僃儔乕僛堚揱巕傪IRT1僾儘儌乕僞乕偱嬱摦偡傞偙偲偑偱偒側偄曄堎懱偺慜曽堚揱妛揑(forward genetic) 僗僋儕乕僯儞僌乮Kim傜丄2019乯丄ILR3偺嫟柶塽捑崀偲塼懱僋儘儅僩僌儔僼傿乕僞儞僨儉幙検暘愅乮Gao傜丄
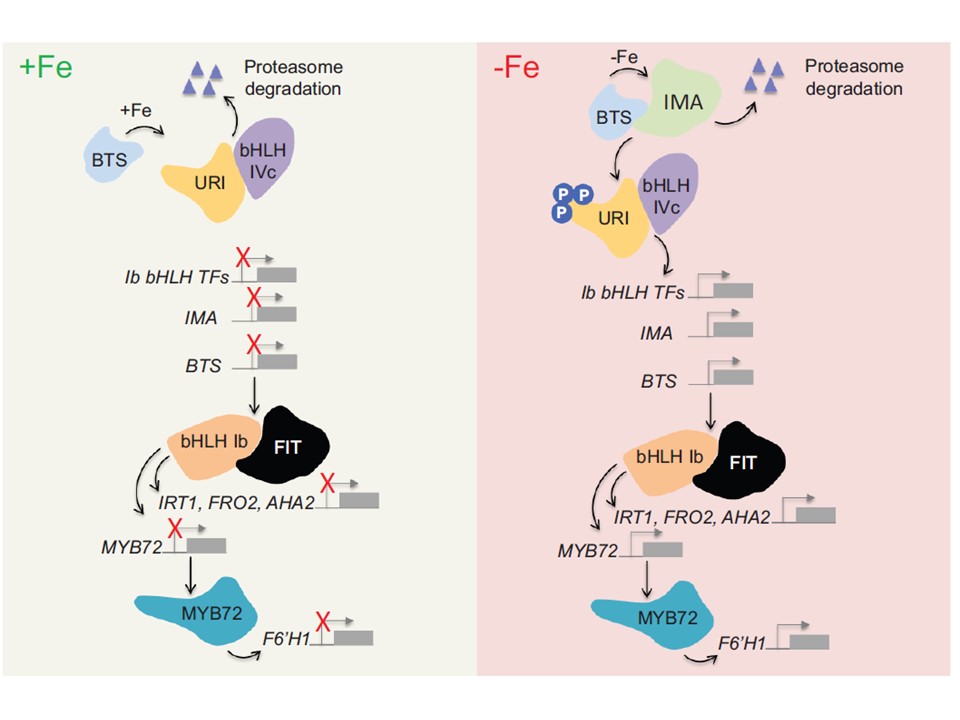
2020乯丄FIT僾儘儌乕僞乕傪塧偲偡傞峺曣儚儞僴僀僽儕僢僪僗僋儕乕僯儞僌乮Lei傜丄2020乯偺3偮偺堎側傞傾僾儘乕僠傪嵦梡偟丄僷僘儖偺儈僢僔儞僌僺乕僗偱偁傞 clade IVb bHLH僞儞僷僋幙 URI 傪摿掕偟偨丅
揝寚朢忦審壓偱URI偼丄揝寚朢忦審壓偱儕儞巁壔偝傟丄僾儘儌乕僞乕偲偺寢崌傪懀恑偡傞廋忺傪庴偗傞丅
偙偺廋忺偼丄偦偺昗揑偺僾儘儌乕僞乕傊偺寢崌傪懀恑偡傞傕偺偱偁傞
(Kim et al.2019乯丅傑偨丄URI僞儞僷僋幙偺嵶朎撪嬊嵼偼丄嵶朎撪僔僌僫儖揱払婡峔偵傛偭偰寛掕偝傟傞丅
URI僞儞僷僋幙偺嵶朎撪嬊嵼偼丄怉暔偺揝偺忬懺偵傛偭偰寛掕偝傟傞丅
揝偺検偑晄懌偡傞偲丄URI偑拞怱拰/撪旂偐傜堏摦偟丄旂幙/昞旂偵堏摦偟丄揝偺媧廂傪懀恑偡傞偙偲偑抦傜傟偰偄傞乮Gao et al.2020乯丅
僒僽僌儖乕僾IVc堚揱巕偲摨條偵丄URI偼揝偵傛偭偰揮幨惂屼偝傟偰偍傜偢丄揝墳摎惈堚揱巕偺僇僞儘僌壔傪栚揑偲偟偨僩儔儞僗僋儕僾僩乕儉夝愅偵偍偄偰丄帠慜偵摨掕偝傟摼側偐偭偨丅
揮幨挷愡偺戙傢傝偵丄儕儞巁壔URI偺儗儀儖偼丄BTS傪夘偟偨僾儘僥傾僜乕儉暘夝偵傛偭偰挷愡偝傟乮Kim傜丄2019乯丄偙偺塣柦偼丄僒僽僌儖乕僾IVc僞儞僷僋幙偵偮偄偰埲慜偵婰弎偝傟偰偄傞偲偍傝偱偁傞乮Long傜丄2010丄Selote傜丄2015乯丅
BTS偼URI偺捈愙偺昗揑偱偁傝丄URI偑弶婜偺揝姶抦乛僔僌僫儖揱払僀儀儞僩偵娭梌偟偰偄傞偙偲傪帵偟偰偄傞丅URI偼IMA1傗IMA2偺敪尰傕捈愙揑偵妶惈壔偡傞乮恾1乯丅
丂丂
丂丂
亙岝偲揝偺僔僌僫儖揱払宱楬偺HY5偵偍偗傞廂澥亜
丂丂
揝偺廀梫偼憡懳揑側怉暔偺惉挿懍搙偵埶懚偟丄怉暔偺惉挿夁掱偱曄壔偡傞乮Grusak 1995,丂Schmidt and F丯uhner 1998)丅
梩傗庬巕側偳偺媧廂婍姱偺揝廀梫傪崻偵揱偊丄搚忞偐傜偺揝偺媧廂傪挷惍偡傞偨傔偵偼僔僌僫儖揱払宱楬偑昁梫偱偁傞丅
FIT傪偼偠傔偲偡傞懡偔偺揝庢傝崬傒堚揱巕偺敪尰偼崻偵尷傜傟傞偨傔丄挿嫍棧捠怣庤抜偑昁梫偱偁傞偙偲偑嫮挷偝傟偰偒偨丅
偙偺傛偆側崻偐傜抧忋晹傊偺僔僌僫儖揱払傪帵偡弶婜偺徹嫆偼丄揝傪峔惉揑偵寚偔怉暔懱偲偺憡屳愙偓栘幚尡偐傜摼傜傟偨丅
僄儞僪僂偺揝寚朢惈峔惉揑撍慠曄堎懱brz偲dgl傪梡偄偨愙偓栘幚尡偐傜摼傜傟偨傕偺偱丄梩偵桼棃偡傞場巕偑撍慠曄堎懱昞尰宆偺尨場偱偁傞偙偲偑帵嵈偝傟偨乮Welch and LaRue 1990, Grusak and Pezeshgi 1996乯丅
偝傜偵丄崻宯偺堦晹傪揝偺側偄梴塼偵偝傜偟丄巆傝偺敿暘傪揝偵傾僋僙僗偝偣傞崻暘偗幚尡偱偼丄崻偺揝媧廂傪挷惍偡傞偨傔偵丄嬊強揑偍傛傃慡恎揑側僔僌僫儖偑摑崌偝傟傞偲偄偆奣擮傪巟帩偟偨(Schmidt and Schikora 2001, Vert et al. 2003乯丅
梩偺揝偺忬懺偑偳偺傛偆偵崻偵揱偊傜傟傞偐偲偄偆媈栤偼丄挿擭偺壽戣偱偁偭偨丅
岝壔妛宯偼揝傪娷傓僞儞僷僋幙傪朙晉偵娷傫偱偍傝丄怉暔偵偍偗傞揝偺嵟傕嫮椡側媧廂尮(僔儞僋)偱偁傞丅
揝寚朢偺姶抦偼梩偺堐娗懇宯偱嵟傕懍偔婲偙傞傛偆偱偁傝乮Khan et al.2018乯丄僔儞僋婍姱偵偍偗傞揝寚朢偺姶抦偲慡恎揑側僔僌僫儖揱払偑崻偺揝媧廂偺惂屼偵偍偗傞弶婜梫場傪峔惉偡傞偙偲偑帵嵈偝傟偨丅
偟偨偑偭偰丄埲壓偺傛偆偵峫偊傞偙偲偼懨摉偱偁傞丅
偍偦傜偔懠偺嵶朎晹暘偵偍偗傞揝偺姶抦偵壛偊丄梩椢懱偱揝傪姶抦偟偰偄傞偲峫偊傜傟傞乮Bienfait et al.1987乯丅
偙偺僔僫儕僆偱偼丄崻偵傛傞揝偺庢傝崬傒偼丄梩椢懱媡峴惈僔僌僫儖偲揝偺忬懺偺慻怐娫揱払偺椉曽偵傛偭偰巟攝偝傟傞丅
揝偲岝僔僌僫儕儞僌傪儕儞僋偝偣傞偙偲偼丄揝偺廀梫偲崻偺庢傝崬傒傪摑崌偡傞偨傔偺榑棟揑側愙懕偱偁傞偲巚傢傟丄偙偺榞慻傒偼丄嵟嬤丄僩儅僩怉暔偵偍偗傞ELONGATED HYPOCOTYL 5乮HY5乯偺僼傿僩僋儘乕儉B埶懚惈拁愊偲揝僔僌僫儕儞僌偺娫偺梊婜偣偸愙懕傪柧傜偐偵偟偨怴偟偄抦尒偵傛偭偰僒億乕僩偝傟偰偄傞乮Guo et al.丄2021擭乯丅
HY5偼梩偐傜崻傊堏摦偡傞偙偲偑偱偒傞墫婎惈儘僀僔儞僕僢僷乕乮bZIP乯揮幨場巕偱偁傞乮Chen et al.丂2016).
揝寚朢忦審壓偱丄HY5偼僩儅僩偺FIT僆儖僜儘僌偱偁傞SlFER偺僾儘儌乕僞乕偵寢崌偟丄揝庢傝崬傒堚揱巕偺敪尰傪桿摫偟丄偦偺寢壥揝偺庢傝崬傒傪桿摫偡傞偙偲偑傢偐偭偨丅
HY5偼梩偲崻偺椉曽偵懚嵼偡傞偑丄憡屳愙偓栘幚尡偵傛傝丄僔儏乕僩桼棃偺HY5偑FER偺妶惈壔偵廳梫偱偁傞偙偲偑帵偝傟偨丅
偙偺傛偆偵丄HY5偼僔儞僋偐傜僜乕僗傊偺揝偺忬懺傪揱偊傞丄懸朷偺儌僶僀儖僔僌僫儖偱偁傞丅
偟偐偟丄HY5偵偳偺傛偆側摿堎惈偑梌偊傜傟偰偄傞偐偼傑偩晄柧偱偁傞丅
HY5偼拏慺丄儕儞巁丄摵偺妉摼偵娭梌偟偰偍傝乮Zhang et al. 2014, Chen et al. 2016, Sakuraba et al. 2018乯丄HY5偑懠偺惂屼場巕偲嫤挷偟偰揝偺庢傝崬傒傪巟攝偟偰偄傞壜擻惈偑帵嵈偝傟偨丅
嫽枴怺偄偙偲偵丄HY5偺夁忚敪尰偼丄怉暔偑揝寚朢傪宱尡偟偰偄傞応崌偵偺傒丄揝寚朢斀墳傪桿摫偟偨丅偙偺娤嶡偼丄偙傟傑偱丄FER傗FIT偺夁忚敪尰偱娤嶡偝傟偨偙偲偲帡偰偄傞丅
揝偑廫暘側忦審壓偱偼尠挊側岠壥傪帵偝側偄偙偲偼(Brumbarova and Bauer)
(Brumbarova and Bauer 2005, Meiser et al. 2011)嬌傔偰廳梫側偙偲偱偁傞丅
揝墳摎惈堚揱巕偺懡偔偼丄僾儘儌乕僞乕偵HY5偲偺寢崌傪壜擻偵偡傞僐儞僙儞僒僗儌僠乕僼傪桳偟偰偍傝丄偙偺帠偼丄HY5偑揝偺僔僌僫儖揱払偵偍偄偰傛傝峀偄栶妱傪扴偭偰偄傞偙偲傪帵嵈偟偰偄傞丅
HY5偑偳偺傛偆偵FER偺揮幨妶惈壔偲偄偆恾幃偵摉偰偼傑傞偺偐丄偦偟偰HY5偑僔儘僀僰僫僘僫偱摨條偺栶妱傪壥偨偡偺偐偳偆偐偼丄傑偩夝柧偝傟偰偄側偄丅
丂丂丂
丂丂
<揝傪庢傝崬傓曗彆揑丒惓摑揑僔僗僥儉偑揝偺桝憲傪僆乕僈僫僀僘偡傞>
丂
僔儘僀僰僫僘僫偱偼丄崅恊榓惈僩儔儞僗億乕僞乕IRT1偑崻嵶朎傊偺揝偺桞堦偺怤擖岥傪惂屼偟偰偄傞偲偄偆悇應偼丄怉暔偺揝尋媶偵偍偄偰挿擭偺僪僌儅偱偁偭偨丅
偦偺敪尒埲棃丄偄偔偮偐偺尋媶偵傛傝丄搚忞偐傜偺揝偺庢傝崬傒傪攠夘偡傞IRT1偺廳梫側栶妱偑妋擣偝傟偰偄傞乮Eide et al. 1996丄Vert et al. 2002, 2003丄Barberon et al. 2014丄Dubeaux et al. 2018乯丅
堎側傞僌儖乕僾偵傛偭偰撈棫偟偰幚巤偝傟偨2偮偺尋媶偼丄嵟嬤丄偙偺壖掕偵堎媍傪彞偊丄偙偺傛偆側尒夝偼傕偼傗懨摉偱偼側偄丅
IRT1偵埶懚偟側偄曗彆揑側揝偺庢傝崬傒僔僗僥儉偺壜擻惈偼丄儕儞巁寚怉暔偵偍偗傞揝儗儀儖偺忋徃偲偦傟偵敽偆IRT1偺敪尰梷惂偐傜悇應偝傟丄埲慜偐傜壖掕偝傟偰偄偨偑丄屻幰偺忦審壓偱揝偑偳偺傛偆偵崻偵擖傞偐偼丄偙傟傑偱晄柧偱偁偭偨乮Tsai and Schmidt 2017a偵憤愢偁傝乯丅
儕儞巁寚怉暔偵揟宆揑側揝偺庢傝崬傒偺憹壛偼丄irt1曄堎懱偱傕娤嶡偝傟乮Hoehenwarter et al.2016乯丄旕惓婯偺IRT旕埶懚揑側揝偺庢傝崬傒偲偄偆柦戣傪棤晅偗傞傕偺偱偁傞丅
偙偺傛偆偵丄IRT1僪僌儅偼丄儕儞巁墫寚懝偺僕儗儞儅偵傛偭偰庛懱壔偟偨偺偱偁傞丅
偟偐偟丄戙懼暔幙偵娭偡傞妋屌偨傞徹嫆偼尒偮偐偭偰偄側偐偭偨丅
嵟弶偺尋媶偱偼丄僼傿僩僔僨儘僼僅傾僔僗僥儉偵椶帡偟偨曽朄偱崻偐傜暘斿偝傟偨僋儅儕儞傪揝偲暋崌壔偝偣偨屻丄IRT1旕埶懚揑偵庢傝崬傓偙偲傪曬崘偟偨丅偙偺偙偲偼丄僀僱壢怉暔偲旕僀僱壢怉暔偺揝偺庢傝崬傒愴棯偺庡側堘偄偵斀偡傞敪尒偱偁傞丅
廬棃丄僋儅儕儞偼揝傪僉儗乕僩壔偍傛傃娨尦偡傞偙偲偱摦堳偟丄FRO2/IRT1傪夘偟偨揝偺庢傝崬傒偺婎幙偲側傞偲峫偊傜傟偰偄偨丅偙偺僔僗僥儉偼丄棙梡壜擻側揝傪傎偲傫偳娷傑側偄愇奃幙搚忞偱惉挿偡傞擻椡傪晅梌偟偰偄傞偑乮Tsai et al.2018乯丄僋儅儕儞偼楢懕揑偵惗嶻偝傟丄嵟廔揑偵偼崻寳偱暘夝偝傟傞丅
惗懺妛揑側娤揰偐傜丄傛傝峫偊傜傟傞僔僫儕僆偼丄揝傪僉儗乕僩偟偨屻偵暘斿偝傟偨壔崌暔傪嵞愛庢偟丄暋崌懱偐傜揝傪曻弌偟偨屻偵僋儅儕儞傪嵞棙梡偟丄柍撆側僌儕僐僔僪偲偟偰挋憼偡傞偙偲偱偁傞丅傾僌儕僐儞偲偟偰嵞暘斿偝傟傞慜偵丄柍撆側攝摐懱偲偟偰曐懚偝傟傞丅
愴棯-I怉暔偵偍偗傞怉暔桼棃揝僉儗乕僩偺庢傝崬傒僔僗僥儉偼丄埲慜偐傜憐掕偝傟偰偄偨丅偙偺壖愢偼丄怉暔惈僔僨儘僼僅傾乮Phytosiderophores乯偺暘斿暔偺娤嶡偵婎偯偔傕偺偱偁傞丅
僀僱壢怉暔偑暘斿偡傞僼傽僀僩僔僨儘僼僅傾偼丄僀僱壢埲奜偺怉暔偺揝塰梴偵傕峷專偡傞偙偲偑偱偒傞丅
偙偺壖愢偼丄椺偊偽丄僺乕僫僢僣偲僩僂儌儘僐僔偱娤嶡偝傟偨傛偆偵丄僀僱壢怉暔偑暘斿偡傞僼傽僀僩僔僨儘僼僅傾偑旕僀僱壢怉暔偺揝塰梴偵峷專偡傞偲偄偆娤嶡偵婎偯偄偰偄傞乮Xiong et al.乯丅
偙偺偨傃丄(Robe et al. 2021a)偺尋媶偵傛傝丄偦偺傛偆側儊僇僯僘儉偺暘巕揑側徹嫆偑帵偝傟偨丅
f6亴h1曄堎懱偼丄僼僃儖儘僀儖CoA偐傜6亴-僸僪儘僉僔僼僃儖儘僀儖CoA傊偺曄姺偵寚懝偑偁傞丅僗僐億儗僠儞偍傛傃壓棳偺僋儅儕儞偺惗崌惉傪懝側偆乮Schmid et al.2014乯丅惗崌惉宱楬偺偙偺抜奒偑寚懝偟偰偄傞曄堎懱偼丄僋儅儕儞傪慡偔娷傑側偄偨傔 偙傟傜偺壔崌暔偺庢傝崬傒偺壜擻惈傪挷嵏偡傞偨傔偺桪傟偨僣乕儖傪採嫙偡傞丅
摿偵丄f6亴h1曄堎懱怉暔偼丄栰惗宆怉暔偲堦弿偵嵧攟偡傞偲丄僌儕僐僔儖壔偝傟偨僋儅儕儞傪娷傓偙偲偑傢偐偭偨丅偙偺偙偲偼丄妶惈偺偁傞傾僌儕僐儞宆偱暘斿偝傟偨僋儅儕儞偼丄儊儕乕僑乕儔儞僪揑側儕僒僀僋儖夞楬偱嵞暘斿偝傟傞慜偵丄僌儕僐僔儖壔偝傟偰丄僌儕僐僔僪偲偟偰挋憼偝傟偆傞偙偲傪帵嵈偟偰偄傞乮Robe et al.丄2021b)丅
摨偠僌儖乕僾偵傛傞捛愓尋媶偱丄僇僥僐乕儖僋儅儕儞偱偁傞僼儔僉僙僠儞偲宍惉偝傟偨Fe3+暋崌懱偼丄幚嵺偵1:3偺妱崌偱僔儘僀僰僫僘僫偺崻偵庢傝崬傑傟丄irt1曄堎懱傪晹暘揑偵媬嵪偱偒傞偙偲偑幚徹偝傟偨乮Robe et al. 2021a乯丅
偙偺傛偆偵丄愴棯-I怉暔偼丄庛巁惈忦審偱嵟傕傛偔摥偔FRO2/IRT1埶懚惈偺揝庢傝崬傒偲丄僋儅儕儞惗崌惉峺慺S8H偺敪尰傪挷愡偡傞偙偲偱pH忋徃帪偵妶惈壔偡傞IRT1旕埶懚偺僋儅儕儞攠夘惈偺揝庢傝崬傒偲偄偆2庬椶偺揝庢傝崬傒婡峔傪梡偄偰偄傞傛偆偱偁傞乮Tsai and Schmidt 2017b丟Gautam傜2021擭乯丅
偟偐偟丄揝-僋儅儕儞暋崌懱偺庢傝崬傒傪攠夘偡傞僩儔儞僗億乕僞乕偼傑偩摿掕偝傟偰偄側偄丅
偙偙偱徯夘偡傞2偮栚偺尋媶偱偼丄IRT1偺堄奜側婡擻傪曬崘偟偰偄傞乮Quintana et al.2021乯丅irt1曄堎懱偱偼丄揝擹搙偑栰惗宆怉暔偲摨摍偐偦傟埲忋偵崅偄偵傕偐偐傢傜偢丄偙偺揝傪抧忋晹偵堏摦偝偣傞偙偲偑偱偒側偄丅
愴棯-I怉暔偵偍偗傞怉暔桼棃揝僉儗乕僩偺庢傝崬傒僔僗僥儉偼丄埲慜偐傜憐掕偝傟偰偄偨丅椺偊偽僺乕僫僢僣/僩僂儌儘僐僔娫嶌僔僗僥儉偱娤嶡偝傟偨傛偆偵丄僀僱壢怉暔偑暘斿偡傞僼傿僩僔僨儘僼僅傾偑旕僀僱壢怉暔偺揝塰梴偵峷專偱偒傞偲偄偆娤嶡偵婎偯偄偰偄偨乮Xiong et al.2013乯丅
偙偺娤應偼丄埲慜 偵傕側偝傟偰偍傝丄揝偼傾億僾儔僗僩拞偵捑揳偟偰偍傝丄IRT1偑婡擻偟偰偄側偄偲梕堈偵庢傝崬傓偙偲偑偱偒側偄偣偄偱偁傞偲偝傟偰偒偨丅
偙偺寢榑偐傜堩扙偟偰丄挊幰傜偼丄彮側偔偲傕偙偺僾乕儖偺僒僽僼儔僋僔儑儞偑崻偺嵶朎偵庢傝崬傑傟傞偙偲傪帵偟丄IRT1旕埶懚揑側揝偺庢傝崬傒偺奣擮傪巟帩偡傞傕偺偱偁偭偨丅偟偐偟丄irt1怉暔偱偼丄崻偺揝僾乕儖偼宻偵桝憲偝傟側偄丅
偙偺偙偲偼丄IRT1偑揝偺崻傊偺庢傝崬傒偵廳梫側栶妱傪壥偨偟偰偄傞偙偲偵壛偊丄崻偐傜抧忋晹傊偺揝偺桝憲偵傕廳梫側栶妱傪壥偨偟偰偄傞偙偲傪帵嵈偟偰偄傞丅 (Quintana丂傜丄2021擭乯丅
IRT1僞儞僷僋幙偺桝憲偵昁恵側巆婎偱偁傞Ser206偵曄堎傪摫擖偟偨僐儞僗僩儔僋僩偱irt1曄堎懱傪曗偭偨偲偙傠丄崻偐傜僔儏乕僩傊偺揝偺暘攝擻椡傪堦晹夞暅偝偣丄irt1曄堎懱偺僋儘儘僔僗昞尰宆傪夞暅偝偣偨丅偙偺偙偲偐傜丄IRT1偼枹抦偺儊僇僯僘儉偵傛傝丄僔儏乕僩傊偺揝揮棳傪娫愙揑偵巟墖偟偰偄傞偲峫偊傜傟傞丅
嫽枴怺偄偙偲偵丄irt1曄堎懱偺僩儔儞僗僋儕僾僩乕儉専嶕偐傜丄揮幨挷愡場巕MYB72偲MYB10偺敪尰偑丄偦偟偰悇掕忋偺MYB72偺昗揑偱偁傞BETA GLUCOSIDASE 42 (BGLU42) 偺敪尰偑IRT1偺婡擻埶懚揑偱偁傞偙偲偑柧傜偐偵側偭偨丅
MYB72偲MYB10偼丄傾儖僇儕惈搚忞偱偺惗懚偲揝屌掕壔僋儅儕儞偺惗嶻偵廳梫偱偁傝丄偙偺娤嶡寢壥偼場壥娭學偑偁傞傛偆偵巚傢傟傞乮Palmer et al.
2014丄Zamioudis et al. 2014丄Stringlis et al. 2018丄Gautam et al.
2021).
BGLU42偼僋儅儕儞偺扙僌儕僐僔儖壔偵廳梫偱偁傝丄崻寳傊偺暘斿偵昁梫偱偁傞丅
偟偨偑偭偰丄IRT1偺婡擻晄慡偑丄梩偺怺崗側揝晄懌傪曗偆偨傔偵丄揝偲暋崌偟偨僋儅儕儞偺惗崌惉丄暘斿丄偦偟偰偍偦傜偔偼庢傝崬傒傪堷偒婲偙偡偲悇應偡傞偙偲偼枺椡揑偱偁傞丅僋儅儕儞傪夘偟偨揝偺庢傝崬傒偼pH偑崅偄偲偒偺傒婡擻偡傞偨傔丄巁惈忦審壓偱偺揝偺庢傝崬傒偵IRT1偑晄壜寚偱偁傞偙偲傪愢柧偡傞偙偲偼廳梫側偙偲偱偁傞丅
丂
丂
亙旝惗暔偵傛傞揝偺庢傝崬傒傪巟墖偡傞揝偺妉摼僔僗僥儉偺傾僂僩僜乕僔儞僌亜
丂丂
崻偺旝惗暔孮偼丄搚忞拞偺儈僱儔儖塰梴慺偺棙梡壜擻惈傪岦忋偝偣傞忋偱廳梫側栶妱傪壥偨偟丄怉暔偺崻偵桼棃偡傞僔僌僫儖偵墳摎偟偰丄娫愙揑偱偼偁傞偑丄嬠椶昦尨懱偵懳偡傞杊屼傪採嫙偡傞乮Stringlis et al.2019, 2021乯丅
傑偨丄崻柺忢嵼嬠偼丄怉暔偺抧忋晹僗僩儗僗墳摎傪巟攝偟丄桪愭偝偣傞偙偲偑柧傜偐偵側偭偨丅旝惗暔-崻-抧忋晹偺僐儈儏僯働乕僔儑儞夞楬偑怉暔偺僼傿僢僩僱僗偵峷專偟偰偄傞偙偲偑帵嵈偝傟偨(Hou et al. 2021)丅
崻偺惗暔憡偺峔憿偲妶惈偼丄搚忞場巕偲摿偵丄怉暔偺崻偐傜攔弌偝傟傞僗僐億儗僠儞丄僼儔僉僙僠儞丄僒僀僪儗僠儞側偳偺僋儅儕儞椶乮Stringlis et al.2018, 2019,丂Voges et al. 2019丄Harbort et al. 2020丄Hou et al. 2021乯偵巟攝偝傟偰偄傞丅
僋儅儕儞惗嶻偺忈奞偼丄摿偵揝寚朢忦審壓偱崻桼棃旝惗暔僐儈儏僯僥傿乕偺慻惉偺僔僼僩傪堷偒婲偙偟丄偙偺帠偼丄揝壱摥壔僋儅儕儞偑旝惗暔偺崻偺忢嵼嬠偺峔憿偲妶摦偵塭嬁傪梌偊傞偲偄偆壖掕傪巟帩偡傞乮Voges et al.2019乯丅
Voges傜偺尋媶偱偼丄偝傜偵丄僇僥僐乕儖僋儅儕儞偑Pseudomonas strain sp. Root329偺憹怋傪梷惂偟偨偑丄廻庡偵桳塿側Pseudomonas simiae WCS417偺憹怋偼梷惂偟側偐偭偨丅偙偺偙偲偼丄崻偺儅僀僋儘僶僀僆乕儉偵偍偗傞偙偺傛偆側曄壔偑怉暔偵偲偭偰桳棙偵側傞偙偲傪帵嵈偟偰偄傞丅
嵟嬤偺尋媶偱偼丄僼儔僉僙僠儞偼丄偦偺捈愙揑側揝壱摥壔妶惈偵壛偊偰丄僐儉僊偺惗堢巋寖偵昁梫偱偁傞偙偲傪帵偟偨乮Harbort et al.2020乯丅
挊幰傜偼丄僗僐億儗僠儞偍傛傃僼儔僉僙僠儞偺暘斿傪寚懝偟偨曄堎懱偑丄揝惂尷搚忞偵偍偄偰栰惗宆偲偼堎側傞儅僀僋儘僶僀僆乕儉傪峔抸偟偨偙偲傪曬崘偟乮Harbort et al.2020乯丄恖岺愇奃壔搚忞乮Stringlis et al.2018乯偍傛傃恖岺攟抧乮Voges et al.2019乯偵傛傞幚尡偐傜摼傜傟偨埲慜偺娤應傪妋擣偟偨丅
摿偵丄嫟惗惗暔偑懚嵼偡傞偲丄f6亴h1曄堎懱偺僋儘儘僔僗昞尰宆傪媬偆偨傔偵昁梫側僼儔僉僙僠儞偺擹搙偑掅壓偟偨丅偙偺偙偲偼丄僋儅儕儞偑搚忞拞偺擄暘夝惈偺揝僾乕儖傪摦堳偡傞埲忋偺婡擻傪帩偭偰偄傞偙偲傪帵嵈偟偰偄傞乮Harbort et al.2020乯丅
偙偺傛偆偵丄怉暔偺崻偲儅僀僋儘僶僀僆乕儉偲偺娫偺壔妛揑僐儈儏僯働乕僔儑儞偼丄憃曽岦揑偱偁傞傛偆偵巚傢傟傞丅偟偐偟丄怉暔偑儅僀僋儘僶僀僆乕儉偺峔憿傪宍惉偟丄崻偺旝惗暔孮廤偑廻庡偺揝偺妉摼傪夵慞偡傞偲偄偆壖愢傪巟帩偡傞徹嫆偑偁傞堦曽偱丄屻幰偵偮偄偰偼丄偳偺傛偆側儊僇僯僘儉偑巟攝偟偰偄傞偺偐偼傑偩晄柧偱偁傞丅
慞嬍嬠偑暘斿傪懀恑偡傞偩偗偱側偔丄僋儅儕儞偺庢傝崬傒傕懀恑偡傞偙偲偱丄偙偺僔僗僥儉傪傛傝岠棪揑偵偟偰偄傞壜擻惈偑偁傞丅
偝傜偵 傑偨丄昦尨嬠偺斏怋偑梷偊傜傟傞偲丄崻寳偱偺揝偺憟扗愴偑尭傝丄娫愙揑偵揝偺庢傝崬傒偑岦忋偡傞偨傔偵怉暔偺惗堢偑椙偔側傞丅
偝傜偵丄僶僋僥儕傾偺僔僨儘僼僅傾乮siderophore乯傪庢傝崬傓棙揰傕偁傞偲偍傕傢傟傞丅偙偺僔僫儕僆偼丄偝傜側傞挷嵏偑昁梫偱偁傞偑丅
偄偢傟偵偣傛丄揝偺媧廂偼丄忢嵼嬠偺懚嵼壓偱挊偟偔夵慞偝傟傞傛偆偱偁傞丅
旝惗暔僐儈儏僯僥傿偑懚嵼偡傞偲丄揝偺妉摼偑戝暆偵夵慞偝傟傞傛偆偩丅
偙偺傛偆偵丄儅僀僋儘僶僀僆乕儉偺峔憿傪曄偊傞偙偲偼丄傾僂僩僜乕僔儞僌偡傞偙偲偱丄怉暔偺婡擻揑側儗僷乕僩儕乕傪憹傗偡偙偲偵側傝偆傞丅
丂
丂
亙僯僐僠傾僫儈儞丗僶僀僆僼僅乕僥傿僼傿働乕僔儑儞偵偍偗傞乽擇廳偺戙棟恖乿亜
丂
悽奅偺恖岥偺戝晹暘偼怉暔惈怘昳偵埶懚偟偰偄傞丅揝暘晄懌偵傛傞揝寚朢惈昻寣偺儕僗僋偑偁傞丅揝寚朢惈昻寣偼丄悽奅揑偵嵟傕堦斒揑側旝検塰梴慺偺寚朢徢偱偁傝丄悽奅揑偵嵟傕堦斒揑側旝検塰梴慺偺寚朢徢偱偁傝丄悽奅拞偱栺16壄恖偑滊姵偟偰偄傞丅乮de Benoist and Clugston 2002, Camaschella 2019乯丅
揝偺塰梴幐挷傪夞旔偟丄岞廜塹惗傪岦忋偝偣傞偨傔偵丄夁嫀悢廫擭偵傢偨傝憡摉側搘椡偑暐傢傟偰偒偨丅揝偺娷桳検偑崅偄昳庬偺奐敪偼丄僶僀僆僼僅乕僥僀僼傿働乕僔儑儞偲屇偽傟傞丅
偙偺奣擮偼丄擾妛揑庤朄傗揱摑揑側堢庬偐傜丄偝傑偞傑側堚揱妛揑傾僾儘乕僠
(Connorton and Balk 2019)傑偱傪堄枴偟偰偄傞丅
揝偺僶僀僆傾儀僀儔價儕僥傿丄偡側傢偪丄揝偺媧廂検偼丄彫挵偱媧廂偝傟傞揝偺検偱偁傞丅偙傟偼僶僀僆僼僅乕僥僀僼傿働乕僔儑儞偺岠棪傪寛掕偡傞戞擇偺庡梫側梫場偱偁傞丅
懱撪傊偺媧廂検偼怘梡怉暔偵娷傑傟傞揝偺娷桳検偵斾椺偣偢丄僼傿僠儞巁墫丄僞儞僯儞丄億儕僼僃僲乕儖側偳丄怘昳偵娷傑傟傞偄偔偮偐偺壔崌暔偵傛偭偰揝偺媧廂偑慾奞偝傟丄岠壥偑憡嶦偝傟傞丅
堦斒偵丄怉暔惈怘昳偺旕僿儉揝偼丄摦暔惈怘昳偺僿儉揝傛傝傕媧廂棪偑掅偄丅
摦暔惈怘昳偺旕僿儉揝偼丄摦暔惈怘昳偺僿儉揝偵斾傋丄媧廂棪偑掅偄丅旕僿儉揝偼丄偦偺傎偲傫偳偑晄梟惈偺戞擇揝偺宍偱懚嵼偟丄廫擇巜挵偺僠僩僋儘乕儉偱娨尦偡傞昁梫偑偁傞丅
擇壙偺嬥懏僀僆儞桝憲懱1 (Dcytb) 傪夘偟偰庢傝崬傑傟傞慜偵丄廫擇巜挵僠僩僋儘乕儉b (Dcytb) 偵傛傞娨尦偑昁梫偱偁傞丅
嬥懏僀僆儞桝憲懱1(DMT1)傪夘偟偰庢傝崬傑傟傞丅偟偐偟丄Dcytb僲僢僋傾僂僩儅僂僗偼揝偵娭偡傞摿掕偺昞尰宆傪帵偝側偐偭偨偙偲偐傜丄揝偺暿偺怤擖宱楬丄偍偦傜偔壗傜偐偺怘塧惈場巕偵傛偭偰僉儗乕僩偝傟偰偄傞偙偲偑帵嵈偝傟偨乮Gunshin et al.2005乯丅
嵟嬤偺尋媶偱偼丄偙偺傛偆側揝偺戙懼宱楬偺婎慴偲側傞儊僇僯僘儉偵岝偑摉偰傜傟丄僶僀僆 僼僅乕僥傿僼傿働乕僔儑儞偵偍偗傞僯僐僠傾僫儈儞乮NA乯偺摿暿側栶妱偑柧傜偐偵側偭偨丅
(NA)偑僶僀僆僼僅乕僥傿僼傿働乕僔儑儞偺傾僾儘乕僠偵偍偄偰摿偵廳梫側栶妱傪壥偨偡偙偲偑柧傜偐偵側偭偨乮Murata et al.2021).
NA偼丄NA-auxotroph偺僋儘儘僱儖僶曄堎懱偺峔惉揑側揝寢崌徢偺昞尰宆傪尦偵栠偡乽惓忢壔場巕乿偲偟偰嵟弶偵曬崘偝傟偨旕僞儞僷僋幙惈傾儈僲巁偱丄崅摍怉暔偵晛曊揑偵懚嵼偟丄揝傗摵側偳偺嬥懏偺曔懆偲堏摦偵娭梌偡傞嬥懏僉儗乕僩嵻偱偁傞乮Poch丩azka and Scholz 1984乯丅
僀僱壢偺怉暔偱偼儉僊僱巁宯偺僼傿僩僔僨儘僼僅傾乮phytosiderophore乯偺慜嬱懱偲偟偰婡擻偟偰偄傞丅
僋儘儘僱儖僶曄堎懱偺僋儘乕僯儞僌偵傛傝丄NA崌惉峺慺偺1墫婎偑曄壔偟偰偄傞偙偲偑敾柧偟偨丅乮NAS; Ling et al.1999乯丅
偙偺峺慺偼揝偺儂儊僆僗僞僔僗偵偍偗傞偦偺儐僯乕僋側栶妱偐傜丄NA偼揝偺僶僀僆僼傿儖僩僼傿働乕僔儑儞偺昿斏側僞乕僎僢僩偱偁偭偨丅
NAS偺堎強惈敪尰偼丄嶌暔偺揝娷桳検傪憹壛偝偣乮Nozoye 2018乯丄傑偨丄
儅僂僗丄僯儚僩儕丄Caco-2嵶朎偱昻寣徢忬傪夞暅偝偣偨(Zheng et al. 2010, Lee et al. 2012, Beasley et al. 2020乯丅
NA僉儗乕僩揝偼惗暔妛揑棙梡擻偑崅偄偙偲偑傢偐傞丅
偟偐偟丄NA傪夁忚敪尰偝偣偨怉暔偱揝偺棙梡棪偑岦忋偟偨棟桼偼丄傎偲傫偳夝柧偝傟偰偄側偄丅
NA偼丄惗暔妛揑棙梡擻偺崅偄Fe2+僀僆儞偲晄梟惈偺Fe3+僀僆儞偺椉曽傪僉儗乕僩偡傞乮von Wir丩en et al. 1999乯丅
偙偺摿惈偼丄惗暔妛揑嫮壔嵻偲偟偰偺栶妱傪壥偨偡忋偱廳梫側梫慺偱偁傞偲巚傢傟傞丅懞揷傜(2021)偼丄偙偺摿惈偵傛傝丄揝-NA暋崌懱偑丄捠忢偺旕僿儉揝偺庢傝崬傒宱楬傪塈夞偡傞偙偲傪柧傜偐偵偟偨丅偦偺戙傢傝丄Fe-NA偼丄僾儘僩儞寢崌宆傾儈僲巁僩儔儞僗億乕僞乕SLC36A1傪夘偟偰挵撪偱岠棪揑偵媧廂偝傟丄枌娧捠宆揝桝憲懱僼僃儘億儖僥傿儞1傪捠偠偰挵娗嵶朎偐傜寣塼拞偵攔弌偝傟傞丅
挵娗嵶朎偺偳偙偱偳偺傛偆偵NA偐傜Fe2+偑梀棧偝傟傞偐偼晄柧偱偁傞丅
懞揷傜偺尋媶偵傛傝丄怉暔懱撪偺NA傪憹傗偡偙偲偼丄揝偺娷桳検傪憹傗偡偨傔偺岠棪揑側愴棯偱偁傞偩偗偱側偔丄怉暔惈怘昳偵偍偗傞揝偺惗暔妛揑棙梡擻傪僒億乕僩偡傞傕偺偱傕偁傞偙偲偑帵嵈偝傟偨丅
怉暔惈怘昳偵偍偗傞揝偺惗暔妛揑棙梡擻偺岦忋偑婜懸偝傟傞丅
NA偼儐價僉僞僗嶌暔傗栰嵷偵晛曊揑偵懚嵼偡傞偨傔丄摿掕偺嶌暔傗栰嵷偵尷掕偝傟側偄丅
傑偨丄懞揷傜偺寢壥偼丄NAS偺堎強惈敪尰偵傛傞揝嫮壔怉暔偺嶌惢偵壛偊偰丄
NA-Fe僒僾儕儊儞僩偼丄揝寚朢惈昻寣偺儕僗僋傪寉尭偡傞偨傔偺戙懼庤抜偲側傞壜擻惈偑偁傞丅
丂
丂
(寢榑)
怉暔偑偳偺傛偆偵娐嫬曄摦偡傞揝偺棙梡壜擻惈偵懳張偟丄嵶朎撪偺儗儀儖傪堦掕偵曐偮偺偐偵偮偄偰丄夁嫀2丄3擭偺娫偵懡偔偺偙偲偑柧傜偐偵偝傟偨丅
摿偵丄嵟嬤偺尋媶偱丄僀僱偲僔儘僀僰僫僘僫偺娫偵偝傜側傞椶帡惈偑偁傞偙偲偑柧傜偐偵側傝丄2偮偺揝庢傝崬傒愴棯偺嫬奅偑濨枂偵側偭偰偄傞偙偲偑尠挊偱偁傞丅偙傟傜偺僨乕僞傪峫椂偡傞偲丄彮側偔偲傕奣擮揑偵偼丄2偮偺揝妉摼愴棯偼姰慡偵偼攔懠揑偱偼側偄傛偆偱偁傞丅
嵟嬤偺尋媶偐傜丄僀僱傗僔儘僀僰僫僘僫偑嵦梡偟偰偄傞揝偺忬懺偺儌僯僞儕儞僌傗揝寚朢墳摎偺忋棳挷愡婡峔偵娭偡傞抦尒偑摼傜傟偨偑丄條乆側僔僌僫儖偑摑崌偝傟偰摿掕偺墳摎傪桪愭偝偣傞巇慻傒偵偮偄偰偼丄傑偩懡偔偺抦尒偑巆偝傟偰偄傞丅
椺偊偽丄僋儅儕儞偺暘斿傪挷愡偡傞偨傔偵奜晹偺pH忣曬傪揱払偡傞悇掕僔僌僫儖偲丄怉暔偺堎側傞晹埵娫偱揝偺忬懺傪揱払偡傞悇掕僔僗僥儉僔僌僫儖偲偺憡屳嶌梡偼丄傑偩傎偲傫偳暘偐偭偰偄側偄丅
傑偨丄僋儅儕儞偺暘斿偑丄怉暔偺乽彆偗傪媮傔傞乿斀墳偵偳偺傛偆偵娭梌偟丄桳塿側僶僋僥儕傾偺摦堳傗揝偺媧廂傪懀恑偡傞偺偐傕柧傜偐偵偝傟傞偙偲偑懸偨傟偰偄傞丅
偙傟傜偺忣曬傪傕偲偵丄搚忞偐傜偺揝偺庢傝崬傒岠棪偑岦忋偟丄怘梡怉暔懱拞偺揝擹搙偑崅偔側傞傛偆側怉暔偑惗傑傟傞偙偲偑婜懸偝傟傞丅
The walrus and the carpenter were walking close at hand; they
wept like anything to see such quantities of sand: 乪If this were only
cleared away乫, they said, 乪it would be grand!乫
Lewis Carroll
(偙偺帊偺堄枴偑彫惗偵偼暘偐傜側偄偺偱栿偟傑偣傫偱偟偨)
丂
丂
乮恾1偺愢柧乯
URI偼丄揝偺僔僌僫儖揱払僇僗働乕僪偵偍偄偰拞怱揑側埵抲傪愯傔偰偄傞丅URI/bHLH121偼丄ILR3傗懠偺clade IVc bHLH僞儞僷僋幙偲僿僥儘僟僀儅乕傪宍惉偟偰丄壓棳偺昗揑傪惂屼偟偰偄傞丅
揝偑側偄応崌丄URI偼儕儞巁壔偵傛偭偰妶惈壔偝傟乮惵怓偺朅偱帵偡乯丄昗揑偺僾儘儌乕僞乕傊偺寢崌傪懀恑偡傞丅
揝偑懚嵼偡傞偲丄E3儐價僉僠儞儕僈乕僛BTS偑妶惈壔偝傟丄揮幨場巕暋崌懱偺僾儘僥傾僜乕儉暘夝傪拠夘偟偰揝僔僌僫儖傪掆巭偝偣傞乮巼怓偺嶰妏宍偱帵偡乯丅
揝偑側偄応崌丄IMA儁僾僠僪偼BTS偺寢崌偲嫞崌偟丄壓棳偺堚揱巕偺揮幨傪桿摫偡傞丅
FIT偼URI偵傛偭偰娫愙揑偵clade Ib bHLH僞儞僷僋幙偑桿摫偝傟傞偙偲偱惂屼偝傟丄FIT偲僿僥儘僟僀儅乕傪宍惉偟偰巁惈壔乮AHA2乯丄娨尦乮FRO2乯丄揝偺庢傝崬傒乮IRT1乯傪夘偟偰搚忞偐傜偺揝妉摼傪拠夘偡傞堚揱巕偺揮幨傪傾僢僾儗僊儏儗乕僩偟偰偄傞丅傑偨丄FIT/bHLHIb偼丄MYB宆揮幨場巕MYB72偺桿摫傪夘偟偰僋儅儕儞偺惗嶻丒暘斿傪惂屼偟丄崅pH偱偺娨尦丄僉儗乕僩丄揝偺庢傝崬傒偵傛傞揝妉摼傪曗彆偟偰偄傞丅
Fe3+-僋儅儕儞暋崌懱偼丄FRO2偵傛偭偰娨尦偝傟丄曻弌偝傟偨揝偼IRT1偵傛偭偰庢傝崬傑傟傞丅
偁傞偄偼丄Fe3+-僋儅儕儞嶖懱傪偦偺傑傑庢傝崬傓壜擻惈傕偁傞丅揝-僋儅儕儞暋崌懱偺庢傝崬傒偺偨傔偺僩儔儞僗億乕僞乕偼傑偩尒偮偐偭偰偄側偄丅
偙偙偵帵偡儌僨儖偼丄Kim傜乮2019乯丄Lei傜乮2020乯丄Gao傜乮2020乯丄Li傜乮2021乯偺曬崘僨乕僞偵婎偯偔傕偺偱偁傞丅
恾1