揝傪夘偟偨彫朎懱僗僩儗僗偵傛偭偰堷偒婲偙偝傟傞僆乕僩僼傽僕乕偼丄怉暔偵偍偗傞Pi婹夓偺弶婜抜奒偵偍偗傞廳梫側僗僩儗僗墳摎偱偁傞
埲壓偼擔杮恖尋媶幰偵傛傞ER傪夘偟偨揝偲儕儞巁戙幱尋媶偱偁傞丅傑偩弌斉宖嵹偝傟偰偄側偄偑傾僋僙僾僩偝傟偰偄傞傜偟偄偺偱東栿偟偰傒偨丅愄丄搶戝偺椬偺桳攏孾乮敪峺妛乯尋媶幒偱崅寧丒揷懞(妛憿)傜偑敪尒偟偨峈惗暔幙tunicamycin偑ER僗僩儗僗傪婲偙偡偲偄偆偙偲偼抦傜側偐偭偨偺偱偦偺揰偱傕彫惗偵偼偨傔偵側偭偨丅
乮奣梫乯偽偐傝偱側偔乮峫嶡乯傕栿偟偰偍偄偨丅
揝傪夘偟偨彫朎懱僗僩儗僗偵傛偭偰堷偒婲偙偝傟傞僆乕僩僼傽僕乕偼丄怉暔偵偍偗傞Pi婹夓偺弶婜抜奒偵偍偗傞廳梫側僗僩儗僗墳摎偱偁傞
丂丂
媑晲梇巙丄幝嶈戝庽丄媑杮岾婌 (柧帯戝妛擾妛晹乯
丂丂
乮奣梫乯
柍婡儕儞巁乮Pi乯偼怉暔偺惗堢偵晄壜寚偱偁傞丅偟偐偟丄搚忞拞偱偼儕儞巁偑惂尷偝傟傞偙偲偑懡偄丅
偦偺偨傔丄怉暔偼儕儞巁寚朢偵懳墳偡傞偨傔偺偄偔偮偐偺婡峔傪妋棫偟偰偄傞丅
偦偺偆偪偺傂偲偮偵丄枌帀幙偺儕儌僨儕儞僌傗暘夝峺慺偵傛傞僾儔僗僥傿僪DNA偺暘夝偑偁傞丅
偟偐偟丄懠偺暘夝宯偑Pi儕僒僀僋儖偵娭梌偟偰偄傞偐偳偆偐偼丄傑偩晄柧側傑傑偱偁傞丅
僆乕僩僼傽僕乕偼丄嵶朎撪峔惉梫慺傪暘夝偡傞僔僗僥儉偱偁傝丄拏慺丄扽慺丄垷墧側偳堦晹偺塰梴慺偺婹夓忬懺偱偺嵞棙梡偵婑梌偟偰偄傞丅
杮尋媶偱偼丄僆乕僩僼傽僕乕寚懝曄堎懱偱偼丄Pi偑憗婜偵屚妷偟丄Pi婹夓壓偱梩偺惉挿偑挊偟偔慾奞偝傟傞偙偲傪尒弌偟偨丅
Pi屚妷偺弶婜抜奒偱桿摫偝傟傞僆乕僩僼傽僕乕偺庡側壸暔偼丄彫朎懱乮endoplasmic reticulum乯偱偁傝丄ER傪慖戰揑偵暘夝偡傞僆乕僩僼傽僕乕偺堦庬偱偁傞ER-phagy偑丄Pi偺儕僒僀僋儖偵婑梌偡傞偨傔偵Pi婹夓偺弶婜抜奒傊偺墳摎偵娭梌偟偰偄傞偙偲偑帵嵈偝傟偨丅
偙偺ER-phagy偼丄彫朎懱僗僩儗僗墳摎偑寚懝偟偨INOSITOL-REQUIRING ENZYME 1擇廳曄堎懱ire1a ire1b偱梷惂偝傟偨偙偲偐傜丄Pi婹夓弶婜偺ER-phagy偼彫朎懱僗僩儗僗偵傛傝桿摫偝傟傞偲悇應偝傟偨丅
偝傜偵丄揝傪惂尷偟丄帀幙-ROS偺拁愊傪梷惂偡傞偲丄ER-phagy偑梷惂偝傟傞偙偲偑傢偐偭偨丅
嫽枴怺偄偙偲偵丄屻敿偺Pi婹夓偵懳偡傞墳摎偱偁傞枌帀幙儕儌僨儕儞僌偼丄ire1a ire1b 偺弶婜 Pi 屚妷忦審壓偱壛懍偝傟偨丅
崱夞偺尋媶惉壥偵傛傝丄Pi婹夓偵懳偡傞墳摎偵偼弶婜偲屻婜偺2偮偺抜奒偑懚嵼偡傞偙偲偑柧傜偐偵側傝丄彫朎懱僗僩儗僗傪夘偟偨ER-phagy偑弶婜抜奒偱偺Pi儕僒僀僋儖偵娭梌偟丄屻婜抜奒偺壛懍傪梷惂偟偰偄傞偙偲偑帵嵈偝傟偨丅
丂丂丂
丂丂丂
乮峫嶡乯
杮尋媶偱偼丄揝傪夘偟偨彫朎懱僗僩儗僗偵傛傞僆乕僩僼傽僕乕偑丄怉暔偵偍偗傞廳梫側僗僩儗僗墳摎偱偁傞偙偲傪柧傜偐偵偟偨丅
帀幙儕儌僨儕儞僌偵娭傢傞堚揱巕偺敪尰偼丄揮幨場巕偱偁傞PHOSPHATE STARVATION RESPONSE 1 (PHR1) 偵傛偭偰惂屼偝傟偰偄傞 (Pant et al., 2015)丅
Pi廫暘忦審壓偱偼丄PHR1偼SIG1/Pho81/XPR1僪儊僀儞娷桳僞儞僷僋幙偲偺憡屳嶌梡傪夘偟偨僀僲僔僩乕儖儕儞巁偵傛偭偰晄妶惈壔偝傟傞丅
堦曽丄嵶朎偑Pi婹夓忬懺偵側傞偲丄PHR1偼SAP AND MIZ1 DOMAIN-CONTAINING LIGASE1乮SIZ1乯偵傛偭偰SUMO壔偝傟丄偦偺寢壥丄妶惈壔偡傞丅
弶婜偺Pi寚朢忦審壓偱偼丄PSR堚揱巕偺敪尰偼Col傛傝傕atg2-1偍傛傃atg5-1偱崅偐偭偨乮恾1g乯偙偲偐傜丄ER-phagy偵傛偭偰儕僒僀僋儖偝傟偨Pi偑PHR1妶惈傪梷惂偟偰偄傞壜擻惈偑帵嵈偝傟偨丅
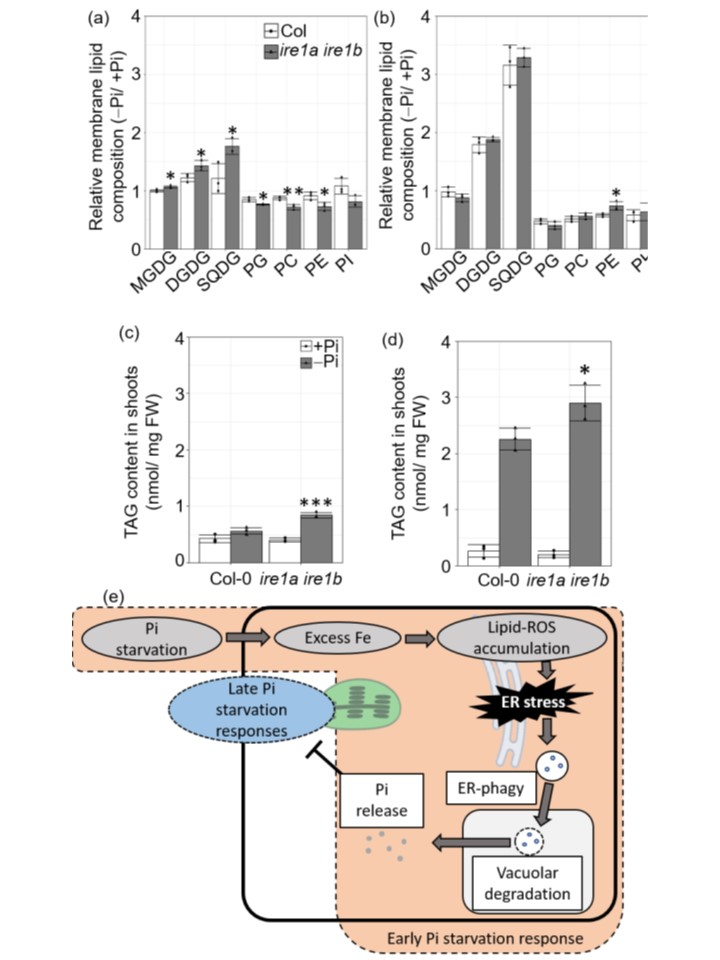
偝傜偵丄ire1a ire1b 曄堎姅偱偼丄弶婜偺Pi寚朢忦審壓偱枌帀幙偺儕儌僨儕儞僌偑懀恑偝傟偨乮恾4a乯丅
ER僗僩儗僗桿摫暔幙偱偁傞僣僯僇儅僀僔儞偵傛傞抁婜娫偺張棟偼丄僔儘僀僰僫僘僫偺儕儞帀幙戙幱僼儔僢僋僗偵塭嬁傪梌偊側偐偭偨丅
偟偨偑偭偰丄ER僗僩儗僗偼偍偦傜偔丄ER-phagy傪庴偗偢偵帀幙慻惉傪捈愙曄挷偝偣傞偙偲偼側偄丅
埲忋偺偙偲偐傜丄弶婜Pi婹夓墳摎偵傛偭偰儕僒僀僋儖偝傟偨Pi偼丄PHR1傪夘偟偨PSR堚揱巕敪尰桿摫傗枌帀幙儕儌僨儕儞僌側偳偺屻婜Pi婹夓墳摎傪梷惂偟偰偄傞偙偲偑傢偐偭偨乮恾4e乯丅
傑偨丄Fer-1偑弶婜Pi寚朢忦審壓偱ER-phagy妶惈傪掅壓偝偣偨偙偲偐傜乮恾3e,f乯丄杮尋媶偱採埬偟偨弶婜Pi婹夓墳摎偼帀幙ROS傪夘偟偨ER僗僩儗僗偵傛傝桿摫偝傟傞偲峫偊傜傟傞丅
堦曽丄屻婜Pi婹夓墳摎偼丄嵶朎撪Pi儗儀儖偺掅壓偵傛偭偰堷偒婲偙偝傟傞丅
怉暔偑挿婜偺Pi婹夓偵偝傜偝傟傞偲丄ER-phagy偵傛偭偰嫙媼偝傟傞Pi偑怉暔偺惉挿偵廫暘偱側偔側傝丄偦偺寢壥丄late Pi starvation墳摎偑桿摫偝傟傞丅
偟偐偟丄屻婜Pi婹夓墳摎偼丄崻栄偺怢挿丄崻偺峔憿偺曄壔丄枌帀幙慻惉偺曄壔側偳偺摦揑曄壔傪堷偒婲偙偡丅
傕偟丄怉暔偑弶婜Pi婹夓墳摎偱偁傞ER-phagy偵傛偭偰Pi傪夞廂偱偒側偗傟偽丄Pi偑嵞嫙媼偝傟偨偲偒偵恦懍偵惓忢側惉挿傪嵞奐偡傞偙偲偑偱偒側偄丅
杮尋媶偼丄怉暔偺Pi婹夓墳摎偑弶婜偲屻婜偺2抜奒偁傞偙偲傪帵偟乮恾4e乯丄怉暔偑帺慠奅偺晄嬒堦側Pi擹搙忦審偵揔墳偡傞偙偲傪壜擻偵偡傞傕偺偱偁傞丅
杮尋媶偱偼丄昪傪Pi寚朢娐嫬偵堏偟偨屻偺梩偺Pi娷検偺帪娫曄壔傪挷傋偨丅昪傪Pi寚朢忦審壓偵堏偟偨屻丄梩偺Pi検偺帪娫曄壔傪挷傋偨乮恾1f乯丅
DAT0偲1偱偼丄Col偲atg曄堎懱偺Pi検偵嵎偼側偐偭偨丅
偙傟偼丄Pi婹夓偺弶婜偵偼傑偩Pi偺僗僩僢僋偑偁傞偲峫偊傜傟丄帺怘嶌梡偺桳柍偵娭傢傜偢Pi検偵嵎偼尒傜傟側偐偭偨丅
DAT4屻偱偼丄Col偲atg曄堎懱偺娫偱Pi検偵嵎偼側偐偭偨丅
彫朎懱僼傽僕乕偵傛偭偰嵞棙梡偝傟傞Pi偼丄偙偺傛偆側抶偄Pi寚朢忬懺偱偼廫暘偱側偄偲峫偊傜傟傞丅
偟偐偟丄DAT2偍傛傃3偱偼丄atg2-1偍傛傃atg5-1偱偼丄Col傛傝傕Pi検偑彮側偔丄偙偺帪揰偱偼丄僆乕僩僼傽僕乕偑Pi儕僒僀僋儖偵廳梫偱偁傞偲偄偆寢榑偑巟帩偝傟偨丅
堦曽丄埲慜偺曬崘偱偼丄Pi寚朢忬懺偺崻偺暘楐慻怐偵偍偗傞僆乕僩僼傽僕乕偼丄Pi儕僒僀僋儖偺儗僗僉儏乕愴棯偱偼側偄壜擻惈偑崅偄偙偲偑帵嵈偝傟偨丅
偙偺僆乕僩僼傽僕乕偼丄崻抂暘楐慻怐乮RAM乯偲崻堏峴乛憗婜怢挿椞堟偱嵶朎宆摿堎揑側PHOSPHATE DEFICIENCY RESPONSE 2-LOW PHOSPHATE RESPONSE1儌僕儏乕儖傪昁梫偲偟偨丅
杮尋媶偱娤嶡偝傟偨僆乕僩僼傽僕乕偺僾儘僙僗偲昞尰宆偼丄暘楐婜椞堟偱偼側偔丄廫暘偵奼戝偟偨屆偄嵶朎偱娤嶡偝傟偨丅偟偨偑偭偰丄Pi婹夓壓偺梩偵偍偗傞僆乕僩僼傽僕乕偺慱偄偼丄崻偵偍偗傞偦傟偲偼堎側傞壜擻惈偑偁傞丅
偙偺栚揑偺徻嵶傪柧傜偐偵偡傞偨傔偵偼丄偝傜側傞夝愅偑昁梫偱偁傞丅
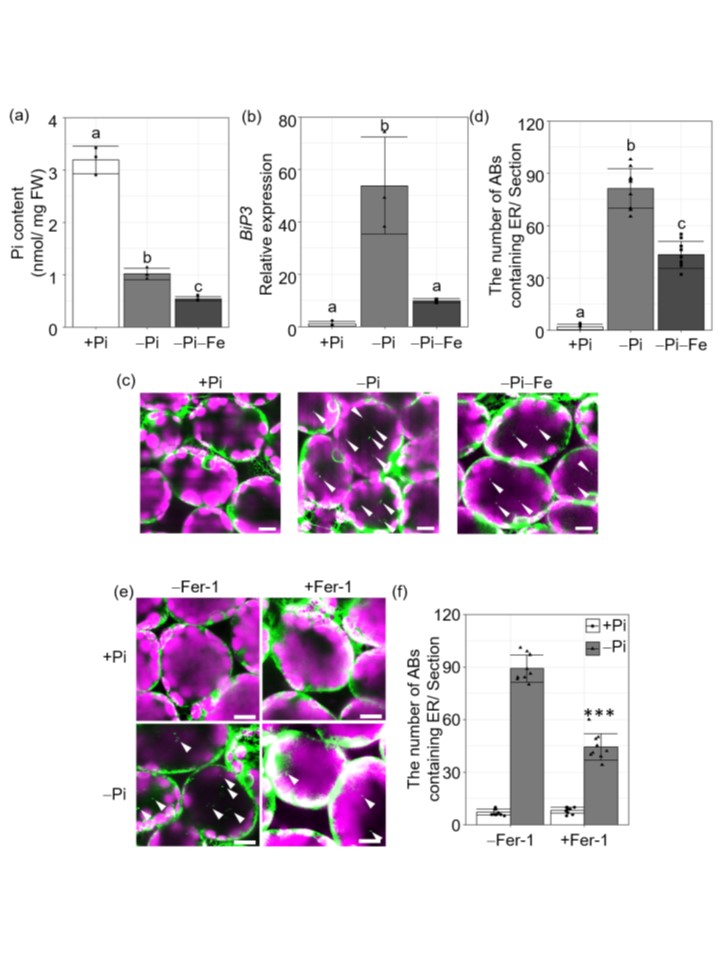
揝偺惂尷偼丄屻婜Pi婹夓壓偱Pi娷検傪憹壛偝偣傞丅偟偐偟丄弶婜偺Pi寚朢忦審壓偱偼丄Fe惂尷偵傛偭偰Pi検偑尭彮偟偨乮恾3a乯丅
Fe惂尷偵傛傝ER-phagy偑梷惂偝傟丄嵶朎撪傊偺Pi曻弌偑梷惂偝傟傞偲峫偊傜傟傞偨傔丄怉暔偼Pi婹夓偺弶婜抜奒偱傕尩偟偄Pi婹夓偵敇偝傟傞偙偲偵側傞丅幚嵺丄-Pi-Fe忦審壓偱偼丄DAT2偵偍偄偰丄-Pi忦審傛傝傕Pi検偑彮側偔丄PSR堚揱巕偺敪尰偑崅偐偭偨乮恾3a乯丅
偙偺傛偆側忬嫷偑挿偔懕偔偲丄PHR1傪夘偟偨Pi庢傝崬傒偺懀恑側偳偺Pi婹夓屻婜墳摎偑憹嫮偝傟傞偲峫偊傜傟傞丅
偙偺傛偆偵丄掅Pi攟抧乮0.1mM Pi乯偱偺Fe惂尷偵傛傝丄Pi娷検偑屻婜偵傾僢僾儗僊儏儗乕僩偝傟傞壜擻惈偑偁傞丅
怉暔偱偼丄僆乕僩僼傽僕乕偑妶惈巁慺偺拁愊傪梷惂偡傞丅偟偨偑偭偰丄僆乕僩僼傽僕乕寚懝曄堎懱偼丄栰惗宆傛傝傕崅偄儗儀儖偱妶惈巁慺傪拁愊偡傞丅偝傜偵丄妶惈巁慺偼僾儘僌儔儉偝傟偨嵶朎巰傪桿摫偡傞丅
変乆偺幚尡偱偼丄atg2-1偍傛傃atg5-1偼丄弶婜偺Pi寚朢忦審偱偼屻婜偺Pi婹夓墳摎偑憹嫮偝傟偨偵傕偐偐傢傜偢丄Pi寚朢忦審壓偱廳搙偺惉挿忈奞傪帵偟偨乮恾1b-d乯乮恾1g乯丅偙傟偼丄妶惈巁慺偺拁愊偵婲場偡傞偲峫偊傜傟傞丅
摦暔嵶朎偱偼丄夁忚側揝偼乽僼僃儘僾僩乕僔僗乿偲屇偽傟傞旕傾億僩乕僔僗惈偺嵶朎巰傪桿摫偡傞丅 偙偺尰徾偱偼丄僼僃儞僩儞斀墳偵傛傝丄嵶朎撪偱Fe2+偲嫟偵Lipid-ROS偑惗惉偝傟傞丅 Lipid-ROS偼丄ER僗僩儗僗偩偗偱側偔丄嵶朎巰傕桿摫偡傞丅
Fe 惂尷偑 Pi 屚妷忦審壓偱惗偠傞惉挿忈奞傪娚榓偡傞傕偆堦偮偺壜擻惈偼丄Fe 惂尷偑嵶朎奜枌偺庡惉暘偱偁傞 PC 偍傛傃 PE 偺嵶朎奜 ROS 偍傛傃僸僪儘僉僔帀朾巁偺拁愊傪梷惂偡傞偲偺娤嶡偐傜愢柧偱偒偨丅
ire1a ire1b 曄堎姅偺枌帀幙偺斾棪偼丄屻敿偺 Pi 屚妷忦審壓偱 Col 偲摨條偱偁偭偨偑丄TAG 娷桳検偼 Col 傛傝傕 ire1a ire1b 曄堎姅偱崅偐偭偨乮恾 4b丄d乯丅Pi婹夓偼TAG偺拁愊傪桿摫偟丄梩偵帀幙揌傪宍惉偡傞丅
僋儔儈僪儌僫僗丒儗僀儞僴儖僨傿偵偍偗傞愭峴尋媶偱偼丄ER僗僩儗僗夁晀惈曄堎懱偱偼丄帀幙揌偺宍惉偑栰惗宆傛傝傕懀恑偝傟傞偙偲偑帵偝傟偰偄傞丅傑偨丄ROS僗僩儗僗偼帀幙揌宍惉傪懀恑偡傞丅
杮尋媶偺寢壥丄Pi婹夓偼梩偺嵶朎偵ER僗僩儗僗偲ROS偺拁愊傪桿摫偡傞偙偲偑柧傜偐偵側偭偨丅
偟偨偑偭偰丄Pi婹夓壓偱偼丄嵶朎撪偺Pi検偺尭彮偱偼側偔丄Pi婹夓偵婲場偡傞ER僗僩儗僗傗ROS拁愊偵傛偭偰TAG偺拁愊偑懀恑偝傟傞壜擻惈偑偁傞丅崱屻丄彫朎懱僗僩儗僗偲帀幙戙幱偺娭學傪偝傜偵徻偟偔挷傋傞偙偲偱丄Pi婹夓壓偱偺TAG拁愊偺儊僇僯僘儉偑柧傜偐偵側傞偲峫偊傜傟傞丅
塰梴婹夓僗僩儗僗偑旕慖戰揑僆乕僩僼傽僕乕傪桿摫偡傞偙偲偼傛偔抦傜傟偰偄傞丅
偟偐偟丄Pi婹夓偵傛傞ER-phagy偼丄ER僗僩儗僗墳摎偵娭傢傞場巕傪棙梡偟偰偄傞傛偆偱偁傞乮恾2d,e乯丅
偝傜偵丄夁忚側徤巁墫偼丄Pi婹夓壓偱RCB傪夘偟偨僋儘儘僼傽僕乕傪桿摫偡傞丅
偝傜側傞妋擣偑昁梫偱偁傞偑丄Pi婹夓偵傛偭偰桿摫偝傟傞僆乕僩僼傽僕乕夁掱偼丄旕慖戰揑偱偼側偔丄慖戰揑偱偁傞傛偆偵巚傢傟傞丅偙偺慖戰惈傪扴偆傾僟僾僞乕僞儞僷僋幙偺扨棧偼丄Pi婹夓偵懳偡傞斀墳慡懱傪姰慡偵棟夝偡傞偨傔偵晄壜寚偱偁傞丅
変乆偼丄揝傪夘偟偨妶惈巁慺偺拁愊偑彫朎懱僗僩儗僗傪桿摫偡傞偙偲傪帵偟偨乮恾3b乯丅
彫朎懱僗僩儗僗偼丄擬僗僩儗僗丄墫僗僩儗僗丄怹摟埑僗僩儗僗偵傛偭偰桿摫偝傟傞丅偙傟傜偺僗僩儗僗偼偡傋偰怉暔儂儖儌儞偱偁傞傾僽僔僕儞巁乮ABA乯偵娭楢偟偰偄傞丅
bZIP17偲S2P偺妶惈壔偼丄ABA僔僌僫儖偺僱僈僥傿僽儗僊儏儗乕僞乕傪僐乕僪偡傞堚揱巕傪傾僢僾儗僊儏儗乕僩偡傞丅
偟偐偟丄ER僗僩儗僗偵懳偡傞ABA偺岠壥傪挷傋偨尋媶偼傎偲傫偳側偄丅
嵟嬤偺尋媶偱偼丄奜場惈ABA偑妶惈巁慺偺拁愊傪桿摫偡傞偙偲偑帵偝傟偨丅
堦曽丄ABA偼僆乕僩僼傽僕乕妶惈傪慾奞偟丄僆乕僩僼傽僕乕偺寚懝偼妶惈巁慺偺拁愊傪桿摫偡傞丅崱屻丄ABA僔僌僫儖傗僆乕僩僼傽僕乕寚懝曄堎懱傪梡偄偨夝愅偵傛傝丄娐嫬僗僩儗僗偑彫朎懱僗僩儗僗斀墳偵梌偊傞塭嬁傪柧傜偐偵偡傞偙偲偑婜懸偝傟傞丅
丂丂
丂丂
丂丂
丂丂
丂丂
乮埲壓恾侾丆俀丆俁丆係偺愢柧乯
丂丂丂
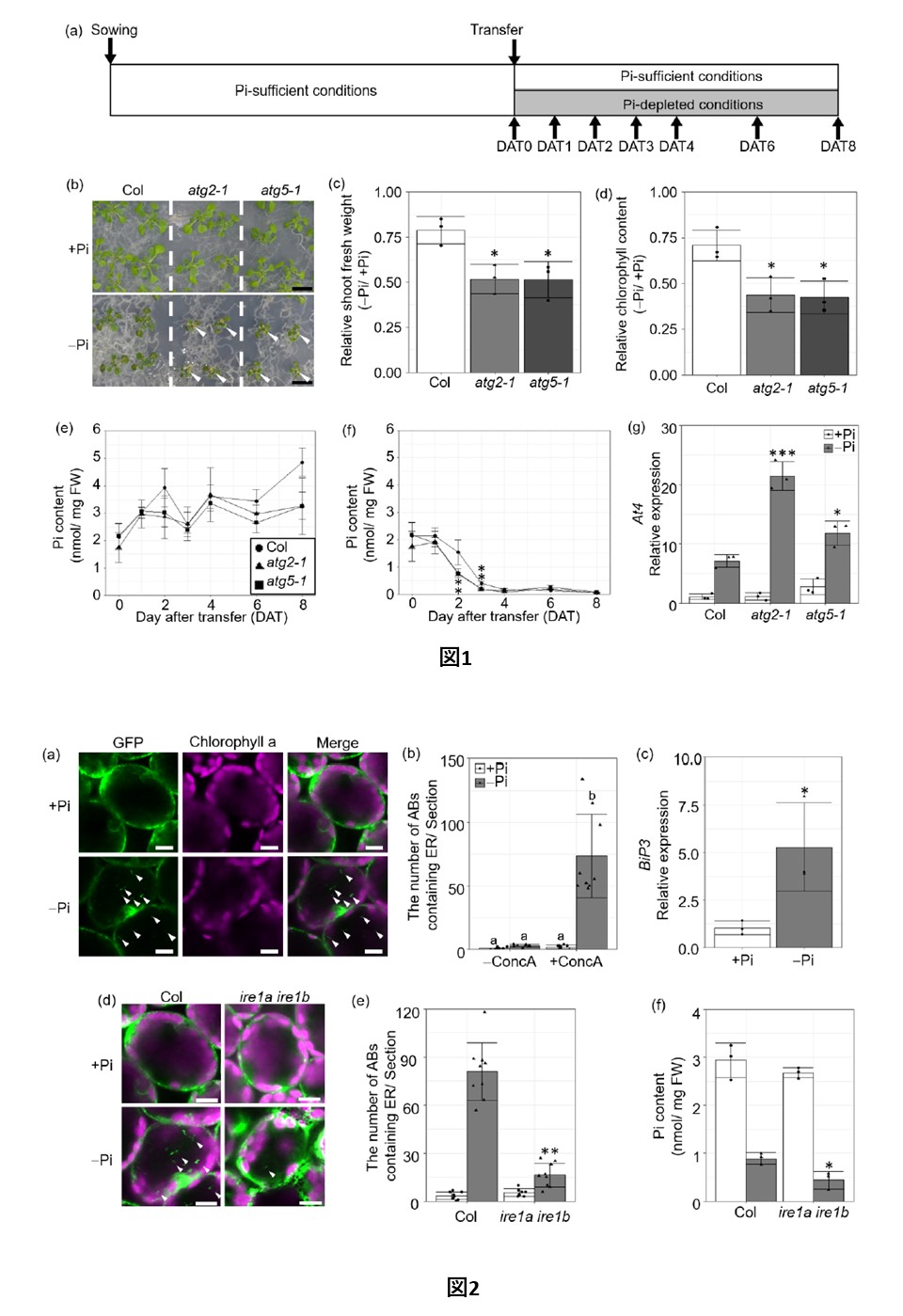
恾1. 僆乕僩僼傽僕乕偼丄Pi寚朢忦審壓偱偺怉暔偺惉挿偵婑梌偡傞丅
(a)丂張棟曽朄恾丅10擔楊偺昪傪Pi晄懌偺攟抧偵攄庬偟丄Pi晄懌傑偨偼-晄懌偺攟抧偵條乆側婜娫堏偟懼偊偨丅怉暔偼丄堏怉屻0丄1丄2丄3丄4丄6偍傛傃8擔栚偵廂妌偝傟偨乮偦傟偧傟DAT0丄DAT1丄DAT2丄DAT3丄DAT4丄DAT6偍傛傃DAT8乯丅
(b)丂DAT8怉暔偺惉挿昞尰宆丅栴報偼僋儘儘僥傿僢僋側梩傪帵偡丅僗働乕儖僶乕亖1cm丅
(c,d)丂 DAT8怉暔偺僔儏乕僩怴慛廳(c)偲僋儘儘僼傿儖検(d)偺-Pi/+Pi斾偺抣丅
(e,f)丂 Pi-sufficient (e) 偍傛傃 -depleted (f) 忦審壓偱偺戞堦梩偍傛傃戞擇梩偺Pi娷検丅(g)丂DAT2怉暔偺戞1梩偍傛傃戞2梩偵偍偗傞At4堚揱巕偺敪尰丅
怉暔偼丄Pi廩懌乮+Pi丟敀朹乯傑偨偼丂-晄懌乮-Pi丟奃朹乯忦審壓偱嵧攟偝傟偨丅僨乕僞偼3楢偺暯嬒亇SD傪昞偡丅*, p < 0.05; ***, p < 0.005 奺忦審壓偱偺僟僱僢僩専掕抣 vs. Col丅FW丄怴慛廳丅
丂丂
恾2. 弶婜Pi婹夓偼丄ER僗僩儗僗傪夘偟偨ER-phagy傪桿摫偡傞丅
(a) 1梩偍傛傃2梩偺惗嵶朎偵偍偗傞ER targeted GFP梈崌僞儞僷僋幙偺LSCM壜帇壔丅梩偼ConcA偲20帪娫僀儞僉儏儀乕僩偝傟偨丅
(b) 帇栰撪乮354.2兪m亊354.2兪m乯偵懚嵼偡傞ER傪娷傓AB偺屄悢丅
(c) DAT2 Col怉暔偺戞1梩偍傛傃戞2梩偵偍偗傞ER僗僩儗僗墳摎堚揱巕BiP3偺敪尰丅
(d) Pi廫暘忦審偲Pi寚朢忦審壓偱偺戞堦梩偍傛傃戞擇梩偺惗嵶朎偵偍偗傞 ER 昗揑 GFP 梈崌僞儞僷僋幙偺 LSCM 偵傛傞壜帇壔丅梩偼ConcA偲20帪娫僀儞僉儏儀乕僩偟偨丅
(e) 帇栰撪乮354.2兪m亊354.2兪m乯偵懚嵼偡傞ER傪娷傓AB偺屄悢丅
(f) DAT2怉暔偺戞1梩偍傛傃戞2梩偵偍偗傞Pi娷桳検丅
怉暔偼Pi廩懌乮+Pi丟敀僶乕乯傑偨偼Pi寚朢乮-Pi丟僌儗乕僶乕乯忦審壓偱惗堢偝偣偨丅栴報偼ER傪娷傓AB傪帵偡丅僗働乕儖僶乕亖10兪m丅GFP寀岝乮椢乯偍傛傃僋儘儘僼傿儖a帺敪寀岝乮儅僛儞僞乯傪帵偡丅
僨乕僞偼3夞乮c,f乯傑偨偼9夞乮b,e乯偺楢悢偺暯嬒亇SD傪昞偡丅
堎側傞暥帤偼Tukey's test (b)偺p < 0.05偱桳堄嵎傪帵偡丅奺忦審壓偱偺+Pi (c) 傑偨偼Col (e,f) 偵懳偡傞Student's t-test 抣偼丄*, p < 0.05; **, p < 0.01 傪帵偡丅
丂丂丂
恾3. 揝傪夘偟偨帀幙-ROS拁愊偼ER-phagy傪桿摫偡傞丅
(a) DAT2怉暔偺戞1梩偍傛傃戞2梩偵偍偗傞Pi娷検丅
(b) DAT2怉暔偺戞1梩偍傛傃戞2梩偵偍偗傞BiP3偺敪尰丅
(c)丂戞1梩偍傛傃戞2梩偺惗嵶朎偵偍偗傞ER昗揑GFP梈崌僞儞僷僋幙偺LSCM偵傛傞壜帇壔丅梩偼ConcA偲20帪娫僀儞僉儏儀乕僩偝傟偨丅
(d) 奺忦審壓偱帇栰乮354.2兪m亊354.2兪m乯撪偵偁傞ER傪娷傓AB偺悢丅
(e) 丂Pi-廫暘傑偨偼Pi寚朢忦審壓偱偺1梩偍傛傃2梩偺惗偒偨梩嵶朎偵偍偗傞ER昗揑GFP梈崌僞儞僷僋幙偺LSCM偵傛傞壜帇壔丅梩偼ConcA偲Fer-1乮+Fer-1乯傑偨偼Fer-1側偟乮-Fer-1乯偲嫟偵20帪娫僀儞僉儏儀乕僩偟偨丅
(f) 帇栰撪乮354.2兪m亊354.2兪m乯偺ER傪娷傓AB偺悢丅栴報偼ER傪娷傓Abs傪帵偡丅僗働乕儖僶乕亖10兪m丅GFP寀岝乮椢乯偍傛傃僋儘儘僼傿儖a帺壠寀岝乮儅僛儞僞乯傪帵偡丅僨乕僞偼3夞乮a,b乯傑偨偼9夞乮d,f乯偺惗暔妛揑暋惢偺暯嬒亇SD傪昞偡丅堎側傞暥帤偼Tukey's test偺p < 0.05偱桳堄嵎傪帵偡(a,b,d)丅***, p < 0.005, 奺忦審壓偱偺僗僠儏乕僨儞僩偺t-専掕抣懳-Fer-1 (f).
丂丂丂丂
恾4. ER-phagy偼枌帀幙偺儕儌僨儕儞僌傪梷惂偡傞丅
(a,b)丂枌帀幙慻惉偺-Pi/+Pi斾偺抣丅DAT3 (a) 偲 DAT8 (b) 偵偍偗傞 Col (White bar) 偲 ire1a ire1b (Gray bar) 偺僔儏乕僩偱偺枌帀幙慻惉偺 -Pi/+Pi 斾偺抣丅
(c,d)丂 DAT3(c)偍傛傃DAT8(d)偵偍偗傞僔儏乕僩偺TAG娷検丅怉暔偼Pi偑廫暘側忦審壓乮+Pi丟敀朹乯傑偨偼寚朢側忦審壓乮-Pi丟奃朹乯偱嵧攟偝傟偨丅
(e)丂弶婜Pi婹夓偵懳偡傞墳摎偺婡峔儌僨儖丅Pi婹夓壓偱偼丄嵶朎撪偵夁忚側Fe偑棳傟崬傒丄ER枌帀幙偑巁壔偝傟傞丅帀幙ROS偺拁愊偵傛傝丄ER-phagy偑桿摫偝傟傞丅ER-phagy偵傛偭偰惗惉偝傟偨Pi偼嵶朎撪偵曻弌偝傟丄Pi婹夓墳摎偺抶墑傪梷惂偡傞丅僨乕僞偼9偮偺惗暔妛揑暋惢偺暯嬒抣亇SD傪昞偡丅奺忦審壓偱偺Col偵懳偡傞Student's t-test偺抣偼丄*, p < 0.05; **, p < 0.01; ***, p < 0.005 偱偁傞丅MGDG, 儌僲僈儔僋僩僔儖僕傾僔儖僌儕僙儘乕儖丟DGDG, 僕僈儔僋僩僔儖僕傾僔儖僌儕僙儘乕儖丟PE, 儂僗僼傽僠僕儖僄僞僲乕儖傾儈儞丟PI, 儂僗僼傽僠僕儖僀僲僔僩乕儖丅
恾1丄恾2
恾3
恾4