鉄とリンの相互依存的な利用が逆行性シグナルを介した光合成を制御する
この論文は鉄欠乏処理でで葉がクロロシスになるときに、P欠乏にしたらその進行が抑えられるという知見に基づいて、その機作を解明するうちに、クロロプラストでの鉄とリンの連携を解明したものである。両者にアスコルビン酸の代謝や輸送が関係しているという、非常に面白い知見を得ている。重要なので、(要旨)と(まえがき)と(考察)を全訳しておいた。
鉄とリンの相互依存的な利用が逆行性シグナルを介した光合成を制御する
Interdependent iron and phosphorus availability controls photosynthesis through retrograde signaling
Hye-In Nam, Zaigham Shahzad, Yanniv Dorone, Sophie Clowez, Kangmei Zhao, Nadia Bouain, Katerina S. Lay-Pruitt, Huikyong Cho, Seung Y. Rhee & Hatem Rouached
NATURE COMMUNICATIONS | (2021) 12:7211 | https://doi.org/10.1038/s41467-021-27548-2 | www.nature.com/naturecommunications
(要旨)
鉄の欠乏は光合成を阻害し、クロロシスを引き起こす。
我々は最近、鉄欠乏によるクロロシスがリンの利用可能性に依存することを明らかにした。
植物がこれらの手がかりをどのように統合してクロロフィル蓄積を制御しているかは不明である。
本論文では、鉄欠乏がリン依存的に光合成遺伝子をダウンレギュレートすることを明らかにした。
また、トランスクリプトミクスとゲノムワイド関連解析により、葉緑体アスコルビン酸トランスポーターをコードするPHT4;4と核内転写因子をコードするbZIP58という二つの遺伝子が、鉄リン欠乏下でステイグリーン表現型につながる光合成遺伝子のダウンレギュレーションを抑制することを突き止めた。
これらの栄養素の共同制限は、bZIP58を必要とするアスコルビン酸生合成遺伝子VTC4の発現を活性化することにより、アスコルビン酸の蓄積を誘導する。
さらに、葉緑体アスコルビン酸輸送が、鉄・リン複合欠乏下での光合成遺伝子のダウンレギュレーションを、活性酸素のホメオスタシスを調節することで防いでいることを明らかにした。
本研究は、光合成を栄養の有無に適応させるための活性酸素を介した葉緑体逆行性シグナル伝達経路を明らかにするものである。
(まえがき)
光合成を行う葉緑体には、核ゲノムにコードされた多くのタンパク質が必要である。
植物は葉緑体から核へのシグナル伝達を厳密に制御しているが、その仕組みは十分に理解されていない。
また、葉緑体には、鉄などの栄養素が十分に蓄積されていることが必要である。
葉に含まれる鉄の最大80%は葉緑体に存在し、電子を供与したり受容したりする能力が電子伝達反応の中心的な役割を担っている。鉄は、すべての電子伝達複合体PSI、PSII、シトクロムb6f複合体、フェレドキシンに含まれ、ヘムや鉄硫黄クラスターなどの補酵素の生合成に必要である。
鉄欠乏(-Fe)環境下で栽培された植物は、クロロシス症状を呈し、光合成が阻害される。
しかし、高リン濃度条件下でも、鉄が十分にあるにもかかわらず、葉が白化することがあり、鉄濃度とクロロフィル蓄積の因果関係が疑問視されている。
さらに、最近、鉄とリン酸の複合欠乏(-Fe-P)条件で栽培したイネは、クロロシスの表現型を示さないことが報告された。
これらの観察から、光合成を制御するための栄養素の利用可能性の相互依存的な効果に関する我々の理解にはギャップがあることが明らかになった。
本研究では、グローバルな遺伝子発現解析とゲノムワイド関連研究(GWAS)を組み合わせて発現量形質座(eQTL)を見出し、鉄およびリン酸の利用可能性に応じてクロロフィル蓄積を制御する制御モジュールを明らかにした。
このモジュールには、VITAMINC4(VTC4)というアスコルビン酸(AsA)合成酵素、PHOSPHATE TRANSPORTER 4;4(PHT4;4) という葉緑体AsAトランスポーター、BASIC LEUCINE-ZIPPER (bZIP58)という推定転写因子が関与していることが判明した。
このモジュールの機能は、光合成を制御する上で、栄養不足が重なる環境下での葉緑体-核間のコミュニケーションの重要性に光を当てている
(考察)
鉄は、植物の成長と発達に重要な役割を果たす微量栄養素である。
自然界では、鉄の供給量が少ないことやpHがアルカリ性であること(例えば、pHが7.0を超えると鉄が利用できなくなる)により、植物が鉄欠乏になることがある。
鉄欠乏はクロロシスを引き起こし、根の生長に影響を与える。
植物が鉄欠乏にどのように反応するか、特に根における鉄の取り込みと植物全体への分布の分子機構については、大きな進展があった。
しかし、植物が光合成を制御するために、どのように鉄欠乏を感知し、反応するのかについては、まだ十分に理解されていない。
鉄は光化学系複合体の必須補酵素であり、葉緑体における電子伝達系のほぼすべての構成要素に存在し、光化学系複合体の必須補酵素である。
本研究では、光合成関連遺伝子(PRG)の発現を制御する葉緑体-核間のシグナル伝達経路と、鉄欠乏環境下での葉緑体形成を制御するシグナル伝達経路を明らかにした。
この経路を制御することで、光合成関連遺伝子(PRG)の発現を制御し、鉄欠乏環境におけるクロロシスを抑制することができる。
この新しく発見された経路を制御することで、植物の光合成活性を向上させながら、養分供給を減らすことができ、圃場での植物の生育に直接的な影響を与えることが期待される。
一般に、鉄欠乏環境でのクロロシスは、葉の鉄欠乏に起因すると考えられてきた。
しかし、本研究で述べたように、FeとPの同時欠乏を受けた植物で最適な光合成が行われることは、この仮定を覆すものである。
鉄欠-リン欠条件下でクロロシスが生じないことを説明するには、2つの可能性がある。
第一に、Fe と P の相互作用がその生物学的利用能に影響を及ぼす可能性があり、Fe と P の同時欠乏は、光合成に利用可能な Fe を増加させる可能性がある。
しかし,我々の方法では,-Fe-P条件で生育した植物の葉において,生物学的利用可能な鉄のレベルが変化することを示唆する証拠を見出すことはできなかった。
第二の可能性は、光合成を制御する鉄欠乏シグナル伝達経路が、鉄の利用可能性によって調節されることである。
この仮説は、我々のグローバルな遺伝子発現解析によって明らかに支持された。その結果、-Fe による 32 PRG のダウンレギュレーションは、Fe と P の同時欠乏によって阻止されることが明らかになった。
また、本研究で発見した PRG 発現の重要な転写調節因子である bZIP58 についても、同様の転写応答が観察された。
鉄とリン酸は相互に作用して根の成長を制御することが知られている。
しかし、光合成を制御するための鉄とリン酸のシグナルのクロストークに関する分子基盤の理解は、まだ不十分である。
そのため、栄養不足の条件下で植物の光合成を改善する鍵となる分子メカニズムを明らかにすることは非常に重要である。
我々は、発現GWASにより、葉緑体PiおよびAsAトランスポーターPHT4;4が鉄欠乏によるクロロシスを制御することを見出した。
この結果は、PHT4;4 の発現が鉄欠乏に影響されないことを示しており、注目される。
その二重輸送活性を考えると、pht4;4変異体が-Fe-P処理下で持続的なクロロシス表現型を示すのは、PiとAsAのどちらの輸送対象が原因であるかが問われることになる。
また、Pi輸送に関しては、他のプラスティドPiトランスポーター(PHT4;3, PHT4;5, PHT4;6)やPiシグナル伝達経路の主要コンポーネント(SPX1とPHR1)を遺伝的に不活性化しても、-Fe +Pまたは-Fe -P条件下で野生型植物と同様の応答を示すことが明らかにされた。
このことは、PHT4;4がAsA輸送活性を介して光合成に影響を与えるという考えを支持する。また、AsAの補給により、野生型植物ではクロロシスの発生を完全に阻止できるが、pht4;4植物では阻止できないことから、この表現型はPHT4;4を介した葉緑体へのAsA輸送に依存することが示唆された。
本研究では、鉄欠乏処理後 52 時間に AsA 量を測定したところ、葉における AsA の蓄積量が有意に減少していることを見出した。
Brassica napus の葉およびテンサイの根では、2-3 週間の鉄欠乏処理に反応して AsA 量が増加することが示され ている。
しかし、シロイヌナズナでは、3-7日の鉄欠乏処理で、新芽のAsA蓄積がわずかに減少することが示された。
鉄欠乏の初期段階において、AsAが一貫して減少していることは興味深い。
1 つの可能性として、AsA の減少は鉄欠乏ストレスの初期徴候の 1 つである可能性がある。
この仮説を支持するのは、塩ストレスに対する初期の反応が、ABI4を介したVTC2の抑制によるAsA生合成の抑制であるという最近の知見である。
AsA生合成を含む、我々が発見したシグナル伝達経路の上流因子を同定することは、今後の重要な方向性であろう。
鉄欠乏はチラコイド膜の電子フラックスの減少を引き起こし、+鉄の条件と比較して活性酸素の増加をもたらす。
いくつかの外来ストレスは、活性酸素の主要な生成場所である葉緑体への活性酸素の蓄積を促進する
近年、活性酸素が逆行性シグナルとして作用し、植物の環境ストレス応答、特に塩ストレス耐性に影響を与えることが提唱されている。
このことから、葉緑体内の活性酸素が鉄欠乏下で逆行性シグナルとして働き、核内PRGの発現を制御しているのではないかという興味深い疑問が生じた。
我々の研究は、-Fe条件下で、活性酸素がプラスチドのシグナルとして働き、bZIP58の抑制を介してPRGの発現とクロロフィル量に影響を与えることを実際に示している。
この効果は、bZIP58を活性酸素誘導性プロモーターの制御下で発現させた場合、より明確になる。
JUNGBRUNNEN1(JUB1)プロモーターの制御下でbZIP58を発現させると、この効果はより明確になる。
JUB1p::bZIP58 を -Fe +P で誘導すると、bzip58 変異体ではクロロフィル蓄積と PRG の発現が促進され、これらの反応は bzip58xpht4;4 二重変異体では悪化することがわかった。
そして、PRGの発現抑制は、葉緑体内の活性酸素が核内転写因子bZIP58の発現を制御することによって行われることが明らかになった。
本研究は、鉄・リン酸複合ストレス下における光合成の制御に関与するシグナル伝達経路を同定し、その妥当性を確認するものである。
この新たなメカニズムは、葉緑体タンパク質(PHT4;4)および核タンパク質(bZIP58)をコードする遺伝子を含み、鉄とリン酸の共欠乏下でコア光合成遺伝子の抑制とそれに伴うクロロシスを防止する。
さらに、bZIP58 が AsA 生合成遺伝子の発現を制御していることを示し、pht4;4 変異株を用いて、AsA の葉緑体への輸送が鉄・リン酸共欠乏下でのクロロシス防止に重要であることを明 らかにした。
また、-Fe条件下では、プラスティドの活性酸素レベルの増加が核でシグナルとして感知され、bZIP58や下流の光合成遺伝子の発現をダウンレギュレートし、クロロシスをもたらすことを明らかにした。
これらの結果は、鉄制限下でのクロロフィル蓄積と光合成に関する基本的な新しい知見を提供するとともに、光合成を栄養条件に適合させる手段として、プラスチド-核間のコミュニケーションを調整するシグナル伝達経路を特定するものである。
(図6の説明)
鉄とリン酸のシグナルを統合し、クロロフィル蓄積と光合成遺伝子を制御するシグナル伝達経路を描いた模式図。
鉄欠乏(-Fe +P)では、核内光合成遺伝子の転写制御の中心であるbZIP58の発現が減少する。
Fe欠乏下でのP制限下(-Fe -P)では、このbZIP58のダウンレギュレーションを防ぎ、VTC4を誘導する。
VTC4の発現誘導にはbZIP58が必要であり、その作用は直接的または間接的である可能性があり、ここでは「X」で表している。
VTC4の発現誘導は、PHT4;4を介し、葉緑体中のアスコルビン酸を増加させることを提案する。
アスコルビン酸量の増加は活性酸素の蓄積を防ぐため、bZIP58とその下流の光合成遺伝子の発現を維持し、「ステイグリーン」表現型につながると推測している。
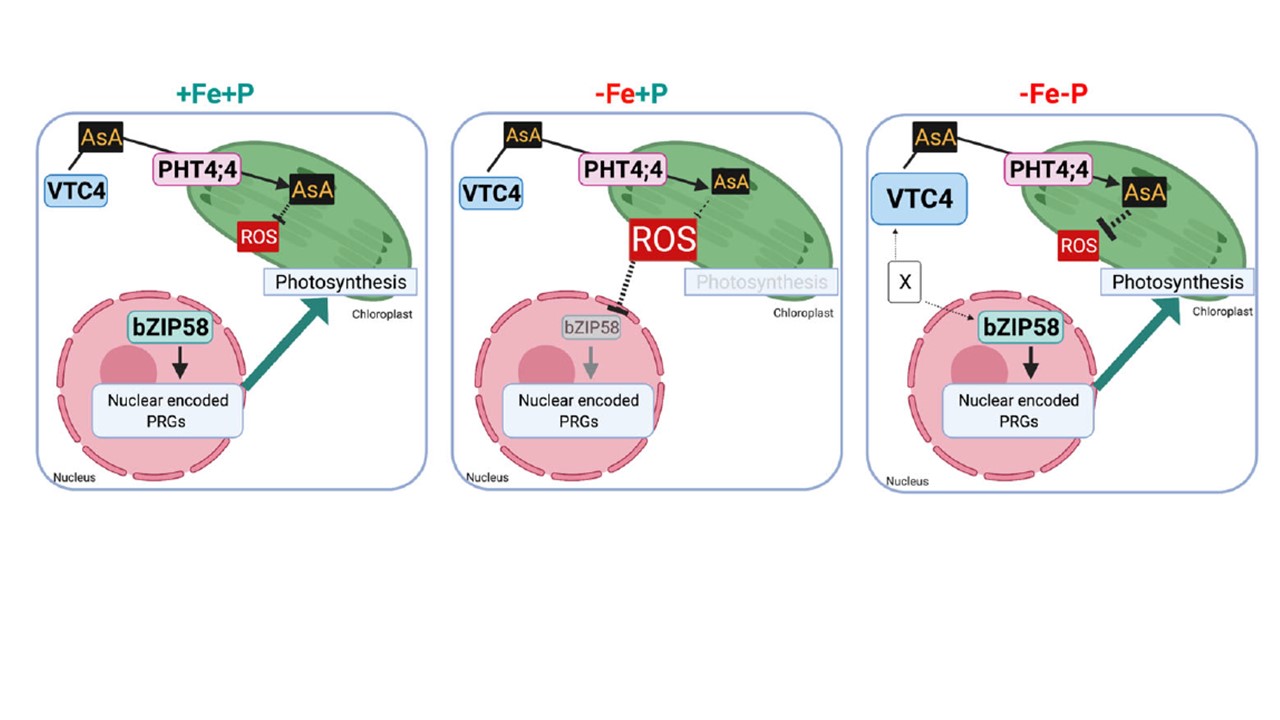
原文の(図6)

原文の(図1)鉄とリン処理の有無によるクロロシスの様相: a浮草, bイネ, cアラビドプシス