リンと鉄のシグナル伝達における相互抑制モジュール
以下の論文は中国人研究者の某君から紹介されたものである。小生はこれまで、鉄(Fe)栄養とリン(P)栄養の関係に関しては、あまり注意を払ってこなかったので、この関係の論文紹介が完全に欠落していた。
日本のリンの研究者は主として土壌の難溶性リンの可溶化・吸収の機構に特化して研究してきているので、リンの体内遺伝子制御機構に関する研究は非常に少ない。ましてやリンと他元素のクロストークの研究は発展途上である。
本論文で紹介している「リンと鉄の体内でのクロストーク」の研究は、非常に新鮮である。中国の研究者頑張っていると思う。そこで、結構時間を取られたのだが、論文の
(要旨)と(まえがき)と)(論議)を全訳しておいた。
リンと鉄のシグナル伝達における相互抑制モジュール
A reciprocal inhibitory module for Pi and iron signaling
Meina Guo, Wenyuan Ruan,Yibo Zhang, Yuxin Zhang, Xueqing Wang, Zhenhui Guo, Long Wang, Tian Zhou, Javier Paz-Ares and Keke Yi
Molecular Plant 15, 138–150, January 3 2022
(要旨)
植物の成長と発達に不可欠な栄養素であるリン(P)と鉄(Fe)は、地殻中に非常に多く存在するにもかかわらず、植物にとって利用しにくい元素であることが多い。植物体内における燐酸塩と鉄の栄養の拮抗作用は、不溶性化合物を形成するため数十年前から注目されていた。しかし、両者の間でのシグナル伝達やホメオスタシスを調節する根本的な分子機構は不明なままである。
本研究では、鉄欠乏誘導性E3リガーゼである鉄センサーHRZが、リン酸(Pi)シグナルの中心的な制御因子であるPHR2と相互作用し、そのリジン残基K319およびK328でのユビキチン化を促して分解する可能性があることをイネで明らかにした。このことと矛盾しないこととして、hrzs変異体は高いPi蓄積表現型を示した。さらに、鉄の不足は、HRZの発現を誘導し、PHR2タンパク質の分解を引き起こすことで、Pi飢餓シグナルを減じることができることを見出した。
一方、イネのPHRはHRZの発現を負に制御し、鉄欠乏応答を調節していることがわかった。したがって、PHR2とHRZは、相互抑制的なモジュールを形成し、Piと鉄シグナルが協調してイネのホメオスタシスを制御している。
これらの結果は、イネにおけるPiと鉄のマスター制御因子が分子的に連携してPiと鉄の利用性を微調整していることを解明したものである。
(まえがき)
リン(P)は作物の生育に不可欠な多量栄養素である。土壌中の全Pは豊富であるが、リン酸(Pi)は、植物が好んで取り込む形態であるにもかかわらず、一般的に土壌への化学的固定が高い上に、土壌中での拡散が遅いため、作物生産の制約要因となっている。安定した高収量を得るために、作物の実際の必要量とは関係なく、毎年リン酸化学肥料が使用されている。その結果、それが環境に対する潜在的な脅威となり、リン肥料生産の再生可能な資源の欠乏を来すという、悪循環がおこっている。そこでPの利用効率を高め、過剰なPi肥料施肥への農業依存を軽減するために、植物体内でのPiシグナルを理解することは、持続可能な社会の実現に向けた大きな課題である。
植物はPi欠乏ストレスに対処するために、Piの獲得と利用効率を高め、Pi飢餓ストレスから守るために、発育中の変化や生化学的な変化などへの異なる応答反応の進化を遂げてきた。これらの反応の制御の中心はMYB-CCドメイン転写因子のサブファミリーであるPHOSPHATE STARVATION RESPONSE (PHRs)として知られ、PHRsは、Pi飢餓に応答する転写産物の大部分を制御している。
シロイヌナズナやイネでは、AtPHR1とOsPHR2の転写レベルは外部からのPi利用性に応答しない。これらのPHRsの活性は、SPX(SYG1、Pho81、XPR1から命名)タンパクによりネガティブに制御される。これらのタンパクはイノシトールピロリン酸シグナル伝達分子(InsPs)存在下で、PHRsと物理的に相互作用することによりPHRsが下流のPi飢餓応答遺伝子を活性化するのを防ぐ。
SPXsによるPHR活性のPiの利用性に応答する知見に比べて植物におけるPHRsのターンオーバーを制御する機構はいまだに不明なままである。
鉄は、土壌中のPと同様に地殻中に豊富に存在する高いにもかかわらず、利用可能性が低いため、植物の成長にとってはしばしば制約的な資源である。
Piは鉄と相互作用して不溶性錯体を形成するので、鉄とリン酸の相互作用の拮抗作用については、植物栄養学においてよく研究されている。Piの飢餓が植物における鉄の蓄積と利用を促進する一方で、高いPi含量は鉄欠乏を引き起こし、葉の鉄分量は十分であるにもかかわらずクロロシスを引き起こすことが示されてきた。
同様に、鉄欠乏は植物にPiの蓄積を促進することができる。最近、トランスクリプトーム解析によってリン酸欠乏ストレスが鉄過剰で誘導される遺伝子をアップレギュレートすることが明らかになった。逆に、鉄欠乏によって誘導される遺伝子は、Pi欠乏ストレスによって抑制された。
これらの観察から、Piの利用可能性の変化は、鉄のホメオスタシスに強く影響し鉄の利用可能性を変化させることがわかる。その逆もまた然りである。
この一連の話において、シロイヌナズナでは、あるシグナル伝達経路が発見されている。これは局所的なPi欠乏反応のシグナル伝達経路であり根端の成長抑制を制御していることが知られている。しかしこれはSPX1-PHR1系とはほぼ無関係である。
イネでは、低リン酸で根の成長は促進されるか影響がないかであるが、局所的なPi欠乏反応を制御するシグナル伝達経路のうち少なくとも1つのコンポーネント(LPRフェロキシダーゼ)が、鉄の蓄積とPi転流に影響を与えることが分かっている。さらに、PHR1がPi-鉄シグナルのクロストークに潜在的に関与している可能性が指摘されている。しかし、Pi欠乏応答の制御因子であるPHRsを中心としたPi-鉄シグナルの相互作用について、その根本的なメカニズムはまだ不明である。
今回、我々はPHRが関与しているPiと鉄のシグナル伝達の間での「制御連携」があることを同定した。
我々は、鉄センサーの可能性が高いHRZn(Hemerythrin motif-containing Really Interesting New Gene- and Zinc finger proteins)がPHR2を直接ユビキチン化し、そのタンパク質量を調節していることを示した。さらに、我々はPHRsがHRZsの発現を間接的に抑制し、その結果相互に抑制する制御モジュールを完成させるということを示した。
このように、Pi欠乏は、HRZsの転写とタンパクレベルを抑制してPHRsの分解を阻止し、Pi欠乏シグナルとPi吸収を促進する一方、鉄欠乏では、HRZs の蓄積を誘導して、PHR2を分解してPiの過剰蓄積を回避し、植物体内での鉄動員能力を高める。
(論議)
過去20年の間に、いくつかの栄養素P、N、Fe、K、Sなどに特異的なシグナル伝達成分の同定に大きな進展があった。それにもかかわらず、最適な成長と生産性の向上に必要な様々な栄養素間での連携した獲得につながるメカニズムの特性解明は遅れている。
最近になってようやくPと窒素の協調獲得に関する研究、硝酸塩センサーとPiセンサーが相互に作用していることが明らかになった。
本研究は、Piと鉄のホメオスタシスの協調のメカニズムについて光を当てたものである。鉄とPiの拮抗的な相互作用は何十年も前から知られていたが、鉄とPiのシグナル伝達を調整する重要な分子機構とホメオスタシスはほとんどわかっていなかった。
イネのPHR2をターゲットとするE3リガーゼを同定しているときに、我々は鉄のシグナル伝達の負の制御因子であるHRZが、PHR2と干渉し、その分解を促進することを見出した。さらに、我々はPiと鉄のホメオスタシスの連携は、HRZsとPHR2が形成する、相互に阻害する性質を示すモジュールで制御されていることを明らかにした。
これまで、PHRsの転写レベルはPiの利用性に応答せず、その転写活性は
はSPXsによってネガティブに制御されていると報告されていた。しかしながらPHR2のタンパク質量は、外部からのPi利用性に応答してよく調整されており、Pi飢餓条件下でも高いタンパク含量を示していた。
そこで我々は、Pi飢餓下でPHR2タンパク質量が増加することは、このストレス下でのHRZsの抑制に強く依存していることを明らかにした。SPXセンサーによる阻害からPHR2が解放される、と同時にPi飢餓下でPHR2の分解を回避するためにHRZsが抑制される、このように、Pi飢餓応答を相乗的に活性化しているのであろう。
HRZは、N末端にHemerythrinモチーフを持つE3ユビキチンリガーゼであり、シロイヌナズナのBRUTUS(BTS)と高い類似性を示している。HRZとBTSは、鉄のホメオスタシスのネガティブレギュレーターとしてよく知られている。hrzs変異株とbts変異株はともに鉄を蓄積し、鉄が不足すると、鉄欠乏に対する耐性が増加する。BTS/HRZsは、鉄のシグナル伝達に関与する転写因子である鉄関連転写因子2(IRO2)、PRI1/2/3(POSITIVE REGULATOR OF IRON HOMEOSTASIS1)、PYE(POPEYE)、FIT(FER-like iron deficiency–induced transcription factor)などの転写因子の分解を促進することが報告されている。
今回、私たちは、イネにおいてHRZsがPHR2の分解も促進し、Pi飢餓のホメオスタシスを制御することを明らかにした。
我々は、HRZ1とHRZ2は、PHR2と相互作用し、それをユビキチン化することを示した。PHR2は、単一の変異体hrz1およびhrz2においてより安定であったことを考えると、PHR2がhrz1 hrz2二重変異体では高度に蓄積されているはずである。しかし、hrz1の稔性が低いために、hrz1 hrz2 二重変異体は得られなかった。hrzs変異体でPHR2が高い集積を示すことと一致して、これらの変異体ではPiが過剰集積し、Pi十分条件下でもPi飢餓シグナルを誘導した。しかし、これらの効果は、PHR2の変異により大部分が抑制された。
さらに、我々は、Piシグナルの中心的な制御因子であるPHRsが、イネにおいてPi欠乏条件下で、低鉄ストレスへの適応に重要な役割を果たすことを示した。
PHRsをノックアウトすると、Pi欠乏ストレスが鉄欠乏症誘導性クロロシスの発生を抑制する能力を減じた。また、このノックアウト変異体は低鉄ストレスに高い感受性を示し、その感受性は主にHRZ2が担っていることを明らかにした。
この知見と一致するように、イネにおいてHRZ2 の過剰発現株は、低鉄ストレスに対して超過敏であった。
またPi飢餓によって鉄の根から茎葉部への移動が促進され、それが変異株phr2では部分的に緩和されるのだが、これがhrz2の変異により回復された。
したがって、鉄欠乏誘導型E3リガーゼによるPHR2の分解は、鉄とPの間の拮抗的な相互作用が、単に化学的性質に起因するのではなく、Piと鉄シグナル伝達の間の密接なシグナル伝達クロストークに起因しているのである。
また、機能的なHRZs-PHRs相互作用がアラビドプシスにも保存されているかどうかを検討した。これまで、BTS/HRZは双子葉植物と単子葉植物の両方で高度に保存されていることが報告されている。
我々は、AtBTSがシロイヌナズナのAtPHR1と相互作用すること、および、Pi飢餓は、PHRsに依存する形でシロイヌナズナでのBTSの発現を抑制することを明らかにした。
ミネラル栄養の移行に重要な経路である維管束周辺において、PHR1およびAtPHL1のノックアウト株が鉄を異常に分布させることが報告されている。
だから、PHRs-HRZsモジュールによる鉄のシグナル伝達とホメオスタシス制御は植物に保存されたメカニズムであるようだ。
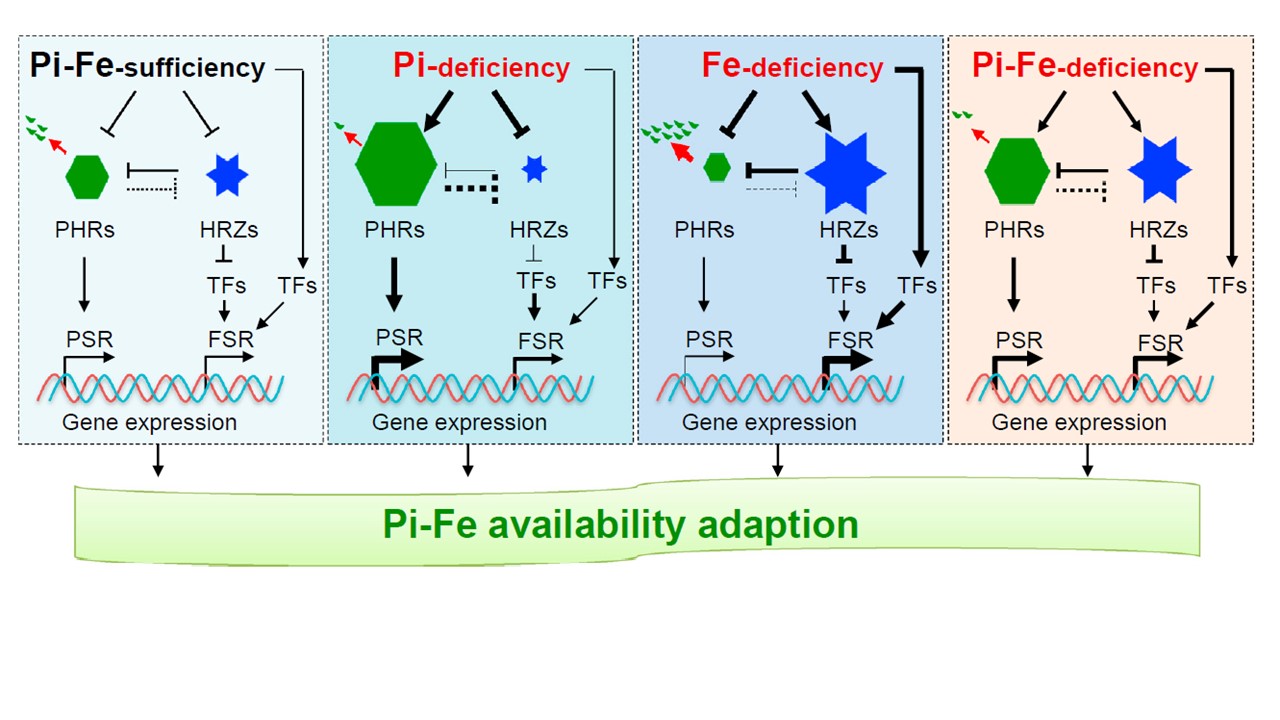
HRZs-PHRsモジュールがどのように連携して鉄/Piのホメオスタシスを制御するのかを示すワーキングモデルを提案する(図6)。
Piと鉄が十分な条件下では、HRZsとPHRsの転写産物とタンパク質は、鉄欠乏応答(FSR)とPSI遺伝子の基礎的発現を可能にするために適切なレベルに維持される。
Piの利用性が低下すると、Pi欠乏によりHRZsの抑制がただちにおこり、
その結果、PHRsのタンパク質量が増加し、さらなるHRZsの発現の阻害がおこる。
PHRsの蓄積の増加と鉄シグナリングの負のレギュレーターHRZsの蓄積の減少は、FSRおよびPSR関連遺伝子発現を活性化し、最終的に鉄蓄積を引き起こし、低Piストレス適応を促進することがわかった。
鉄欠乏状態では、HRZの転写およびタンパク質レベルが増加し、その結果、PHRsの分解が促進される。これは、Piの過負荷を回避するための安全装置である可能性がある。なぜならPiは植物の鉄動員能力を損なう可能性があるからである。鉄とPiの両者ともレベルが低い場合、HRZsとPHRsのタンパク質量は比較的高いレベルに維持され、この2つのストレスに対して、植物はよりよく適応している。したがって、HRZsとPHRsは相互抑制的なモジュールを形成しイネにおけるPiと鉄のシグナル伝達とホメオスタシスを連携させている。
イネにおけるHRZとPHRが制御する鉄とPi栄養間の分子モジュールを理解することは、以下のようなスマートな作物を育成する可能性を秘めている。この作物は(土壌の)異なる鉄とPi利用性のレベルにうまく適応でき、鉄およびPiの獲得量を増加させることができるようになる。つまり石灰質土壌や酸性赤色土壌など、深刻な鉄やPiの利用効率に欠陥がある土壌での利用効率を高めることができる。
この論文では多岐にわたる実験データが示されているので、紙面の都合でそれを紹介することができない。最後の研究成果のモデル図である第6図のみを紹介する。
(図6.の説明)
HRZsとPHRsは相互抑制モジュールを形成し、イネのリン酸・鉄シグナル伝達とホメオスタシスを調整する。
青い六角形はHRZsタンパク質を、緑の六角形はPHRsタンパク質を表している。黒い矢印は正の効果を示す。棒グラフの線は負の効果を、破線は間接的な抑制効果を表す。赤い矢線はタンパク質の分解を示す。また、線の太さは、プラスまたはマイナスの効果の度合いを表している。六角形と六角形の大きさの違いは、PHRsとHRZsのタンパク質または転写レベルの FSRは鉄飢餓応答、PSRはリン酸飢餓応答、TFsは鉄関連転写因子。