葉の中の鉄:化学形態、シグナル伝達、細胞内分布
以下は葉肉細胞を中心にした鉄の代謝制御機構の総説である。これに類似の課題として、
最近発行された「肥料科学第43号」(2021年)に、日本土壌肥料学会賞を受賞した東京農業大学の樋口恭子教授が
光化学系で使われる遷移元素―オオムギ光化学系の鉄利用を中心にー
というタイトルで優れた総説を書いている。
葉の中の鉄:化学形態、シグナル伝達、細胞内分布
Iron in leaves: chemical forms, signalling, and in-cell distribution
Máté Sági-Kazár, Katalin Solymosi, Ádám Solti
Journal of Experimental Botany, Volume 73, Issue 6, 15 March 2022, Pages 1717–1734, https://doi.org/10.1093/jxb/erac030
鉄(Fe)は必須遷移金属である。生体内では酸化還元活性を持つことから、様々な鉄化合物が酸化還元酵素の補酵素として機能している。植物では、光合成装置が最も多くの鉄を必要とする。そのため、光合成装置の補酵素への鉄の供給と取り込みが、葉における鉄代謝の中心的な役割となっている。葉の鉄のホメオスタシスが乱れると、クロロフィルの生合成や光合成装置の構成が損なわれることになる。しかし、ミトコンドリアの機能もまた、鉄に対する大きな需要がある。葉の細胞内で鉄がタンパク質や補酵素に適切に取り込まれ、細胞内の鉄の状態がバランスよく保たれるためには、鉄を感知する能力が必要だが、鉄のホメオスタシスに関連する生理学的プロセスを伝える間接的なシグナルに依存している可能性もある。根における鉄のシグナル伝達については様々な情報が得られているが、葉における鉄の状態の制御については、まだ詳細が明らかになっていない。本総説では、葉における鉄のホメオスタシスについて、化学形態から分配、センシングに至るまで、現在の知見を概観する。
(以下図1,2,3の説明。いささか説明が長いですが、図と照合してください)
図1. 葉における鉄の輸送過程。シロイヌナズナなどの双子葉植物の葉は、主に維管束の根から根への鉄輸送、または葉茎が行うソースシンクの鉄再分配過程を経て、血管系から鉄を受け取る。葉に到達する木部中の鉄は Fe(III)-Cit が主である(1)。木部樹液に由来する鉄カルボキシレートは、細胞内空間やアポプラストに浸潤すると考えられている(2)。アポプラスト空間は弱酸性であるため、鉄カルボキシレートは安定であるが、NAへの配位子交換はまだ明らかにされていない。維管束周囲の柔細胞(3)はYSL1, 3を発現しており、Fe-NAの取り込みも示唆されている。葉肉細胞(4)は主に還元的な鉄の取り込みを行う。FROファミリー酵素は鉄(III)-カルボン酸複合体を利用できるが、葉肉細胞の鉄キレート還元酵素活性の冗長性から、アスコルビン酸を介した鉄(III)-カルボン酸の還元や光還元も関与している可能性が示唆される。木部での鉄の移動に加えて、葉部(5)も鉄の再分配と輸送に関与しており、実際、木部での輸送の方が著しく大きいはずである。茎葉を含む細胞質に満たされた環境では、Fe-NA種が存在し、Feを可溶性に保っていることが示唆される。さらに、長距離の鉄シグナリングに関与する鉄種の輸送も、茎葉輸送経路を利用している可能性がある。鉄の種類に応じた師管への積み下ろし(unloading)の性質はまだ明らかにされていない。細胞間連絡組織(プラスモデスマータ)を介して、細胞質間が連絡してい細胞は、特別な鉄輸送体を必要としないかもしれない。実際、特に葉の発生期や老化開始時にYSLトランスポーターが存在することから、Fe-NA種の輸送もFeの茎葉輸送に重要であることが示されている。
図2. 葉緑体細胞の鉄のホメオスタシス。シロイヌナズナなどの双子葉植物の葉肉細胞は、主に還元に基づく鉄の取り込みを行うが、YSLを介した鉄の取り込みの程度はあまり特徴付けされていない。
二価金属の輸送はIRT型トランスポーターが担っていることが示唆されている。一般に、膜貫通型の輸送は遊離型の鉄であると考えられているが、活性酸素の発生を避けるために、鉄は細胞質で錯体化されることが必要である。
低分子リガンドと複合化した鉄は、細胞内の不安定な鉄プールの一部であると考えられている。この不安定な鉄プールの適切な構成はまだ解明されていない。FROファミリー酵素は、葉緑体にもミトコンドリアにも入り込み、鉄獲得は還元型を基本とする代謝経路に主導されていると考えられている。
ミトコンドリアマトリックスと葉緑体ストロマへの鉄輸送は、オルガネラの二重エンベロープシステムのため、複雑なプロセスである。Vigani et al.(2019)で議論されているように、この器官のタンパク質メンバーの機能的特徴付けは完全とは言い難いものである。
葉緑体のFe獲得では、PIC1、NiCoおよびMFL成分が共働することができるが、ミトコンドリアでは主にMIT1&2を作動させるようだ。葉緑体やミトコンドリアでのFeは、ヘムやFe-Sクラスターへの取り込みに向けられる。
ヘムはフェロキラターゼによって合成されるが、ミトコンドリアでのヘム生合成にはフラタキシンが関与していると考えられている。
鉄硫黄クラスターの生合成には、ミトコンドリアがISCシステム、葉緑体がSUFシステムを作動させている。植物のGRXS(?)は、鉄硫黄クラスターの管理およびアポタンパク質への挿入に関与している。鉄硫黄クラスターの細胞質への輸送には、NEETタンパク質とABCトランスポーターが関与している。光合成および呼吸電子伝達系には大量の鉄が必要であり、葉肉細胞では鉄は主にこれらの過程を担うタンパク質複合体に向けられている。これらのシステムが分解される際に、鉄が再分解され得る。鉄の溶解度を保持し、プラスチドから鉄を排出するために、YSLファミリー輸送が関与していると考えられている。細胞質では、真核生物のCIAシステムが稼働しており、還元硫黄の供給源としてミトコンドリアのISCシステムに一部依存している。葉の細胞の液胞は、鉄の一時的な貯蔵に寄与しているが、鉄の一時的な過剰や鉄の解離を管理するのに役立つ主要な鉄貯蔵は、図3にも示されているフェリチンである。
鉄は潜在的に毒性を持つ元素であるため、細胞の鉄の状態を適切に制御することが重要である。植物細胞の鉄センサーとしては、ヘメリスリンドメインタンパク質BTS(L)が報告されている。
しかし、植物細胞における鉄の状態は複雑であり、小分子やFe-Sを感知する機構を介した複数の感知・制御機構が存在することが示唆されている。
細胞内の鉄ホメオスタシスを制御するエピジェネティックな機構の理解はまだ十分ではないが、Domains Rearranged Methyltransferase (DRM) によるDNAメチル化とヒストン修飾の両方が重要なシグナルであることが示唆されている。
図3. 双子葉植物の様々なプラスティドタイプ、細胞、組織におけるフェリチンの局在。
A: トネリコ(Fraxinus excelsior)の完全に閉じた芽の外葉原基の葉肉細胞のエチオクロロプラスト中のフェリチン(他の詳細はSolymosiら、2012参照)。
B:2週間齢の暗条件で強制栽培されたローズマリー(Rosmarinus officinalis)の葉の葉身柔組織に存在するプラスティドのフェリチン(詳細はBöszörményi et al, 2020を参照)。
C:2週齢の光活性明条件下で成長させたエンドウ(Pisum sativum)の葉のフェリチンを持つ葉緑体。
D: 老化したParthenocissus tricuspidataの葉の葉肉(海綿状柔組織)細胞から採取した老化した葉緑体。矢印はフェリチンを指し、挿入図はプラスティドのフェリチン領域を拡大したもの。電子顕微鏡による試料調製と解析はBöszörményiら(2020)のように行った。スケールバー。1 μm。
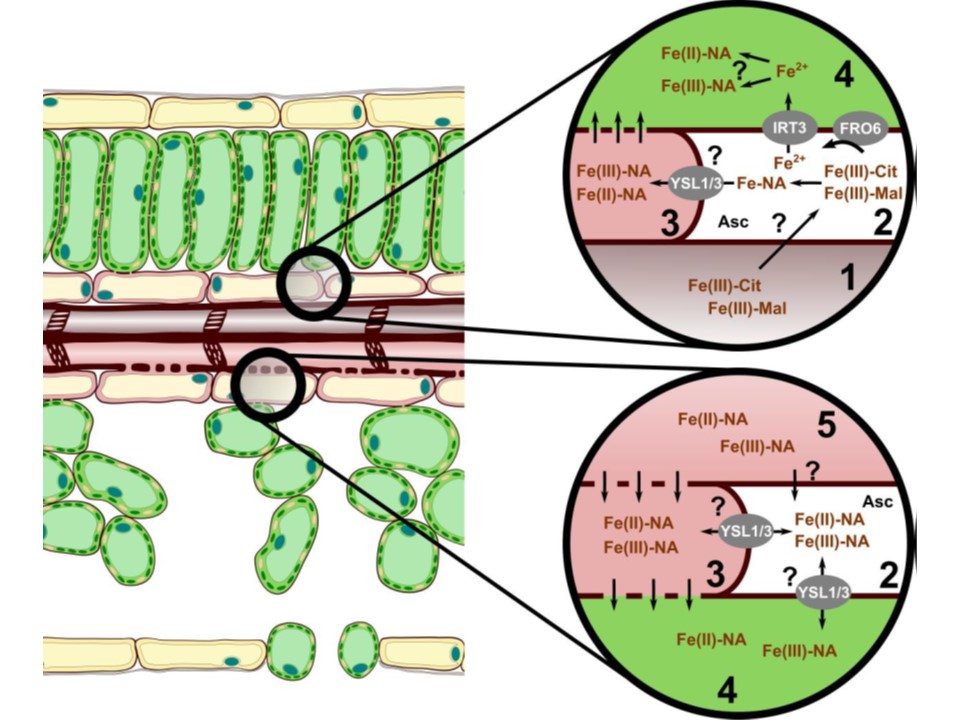
図1
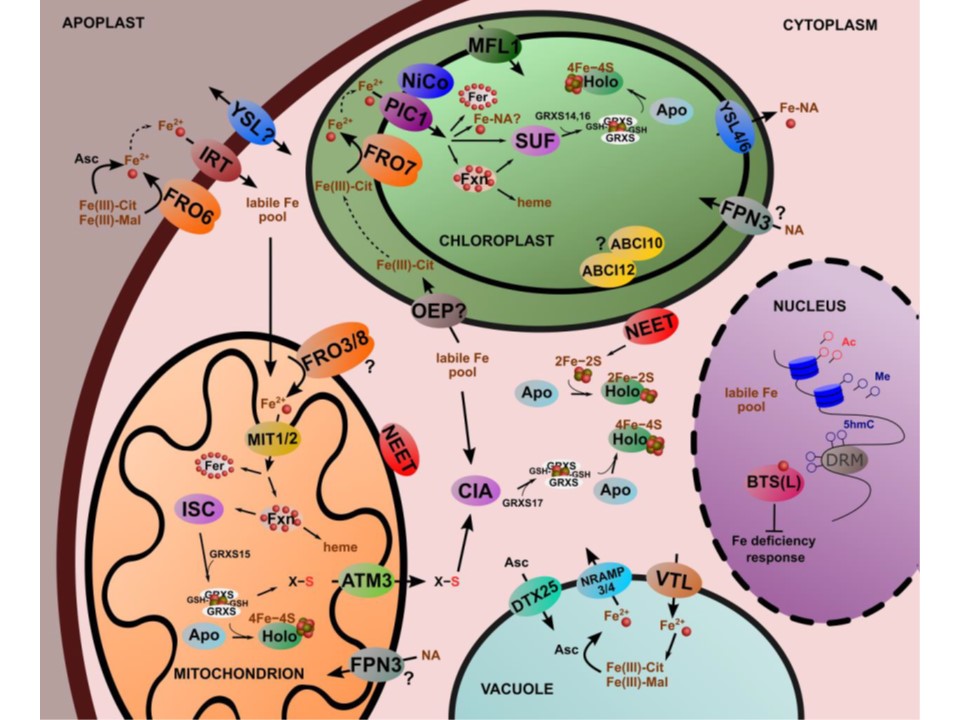
図2
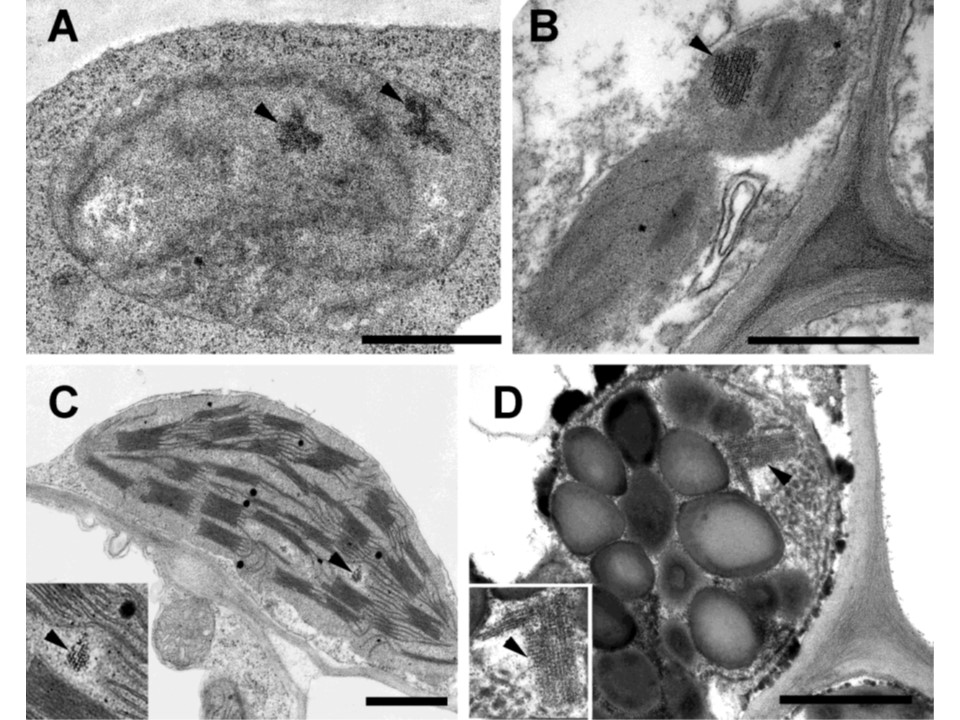
図3