植物の生育に対するアンモニアイオン障害に関する重要論文!:鉄沈積との関連について
私見では、以下の論文は実に卓越した論文であると思う。
筆者の個人的な体験では、小生は半世紀前の若い時から全くの偶然だが、愛媛県農試が開発したハダカムギである「エヒメハダカ」というオオムギの品種を「ムギネ酸研究」の材料に使っていた(これは香川県の篤農家である山下淳二氏を通して入手したものであった)。このオオムギは水耕栽培の時にアンモニア態窒素(NH4+)を唯一の窒素源として育てると根が伸長阻害されて褐変化してほとんど育たなかった。これに対して硝酸態窒素(NO3-)を用いると何の障害もなくよく育った。アンモニア態窒素に対して硝酸態窒素の比を増やしていくと、比例的にアンモニアによる阻害が回復した。また、各種アミノ酸や核酸塩基や高分子タンパクや核酸などを唯一の窒素源にしてもよく育った。
当時、このアンモニア態窒素による生育阻害の理由が小生には全く分からなかった。根圏で何らかの理由でNH3ガスになって、阻害されるのかなと漠然と考えていたにすぎない。もちろん、一方で、水稲の場合はどんな品種でもアンモニア体窒素が最もよく効くことは古くから世界の常識である。
ここで紹介した論文は、図8に示すようにNH4+障害が鉄の過剰集積との関わりで整理されているが、NH4+シグナリング伝達について、まだまだ下流の未解明の経路に関して詰めるところが多く残っている。しかし、小生の従来からのかつての疑問に真正面からから答えるものである。
原著論文には図が8枚もあって紙面の都合上すべてを載せることができないが、すべての図の説明文は訳しておいたので、どういう実験が精緻になされたのかは、想像できると思う。関心をお持ちの方は、ぜひ原著論文を見ていただきたい。
この論文の要旨ばかりでなく論議(discussion)を全文訳した。この論議を読んでいると、この論文の全員中国人の著者らが相当ハイレベルの植物栄養学の知見を理解していることがうかがわれた。中国の浙江大学植物栄養学のレベルがここまで上がっているのかと、少し感慨深かった。
アンモニウムを主要な窒素源とした場合に応答する師管鉄による根の発達の再構築
Phloem iron remodels root development in response to ammonium as the major nitrogen source
Xing Xing Liu, Hai Hua Zhang, Qing Yang Zhu, Jia Yuan Ye, Ya Xin Zhu, Xiang Ting Jing, Wen Xin Du, Miao Zhou , Xian Yong Lin , Shao Jian Zheng & Chong Wei Jin
1 State Key Laboratory of Plant Physiology and Biochemistry, College of Natural Resources and Environmental Science, Zhejiang University, Hangzhou
310058, China. 2 State Key Laboratory of Plant Physiology and Biochemistry, College of Life Science, Zhejiang University, Hangzhou 310058, China.
✉email: jincw@zju.edu.cn
NATURE COMMUNICATIONS | (2022) 13:561 | https://doi.org/10.1038/s41467-022-28261-4 |
(要旨)
植物は硝酸イオンとアンモニウムイオンを主要な窒素(N)源として利用し、それぞれ異なるメカニズムで根の発達に影響を与えている。しかし、これらのイオンが根の発達に関与する正確なシグナル伝達経路は十分に理解されていない。本研究では、シロイヌナズナにおいて、細胞壁に局在するフェロキシダーゼLPR2の破壊または鉄の補給量の減少のいずれかが、窒素源としてのNH4 +に対する初生根の成長阻害を効率的に緩和することを明らかにした。さらに、アンモニウムは硝酸塩と比較して、LPR2 依存的に師管のアポプラストに過剰な鉄を蓄積させることが明らかとなった。このような異常な鉄の蓄積は、その後、活性酸素のバーストから師管に大量のカロースの沈着を引き起こし、師管の機能を低下させた。したがって、アンモニウムはスクロースを十分に生長部に配分できないため、初生根の発達を阻害する。この結果は、環境的な信号に応じた根の形態と師管鉄の関係を結びつけるものである。
(まえがき)略
(実験方法)略
(議論)
NH4 +は植物細胞に吸収された後、直接同化することができるが、NO3 -は植物が利用する前にエネルギーを消費する方法でNH4 +に還元される必要がある。しかしNH4 +が理論的には植物にとって好ましい窒素源であるにもかかわらず、それが唯一または優勢な窒素源である場合、初生根の成長を阻害するという非常に不可解な問題がある。本研究は、初生根 の NH4 + 感受性表現型の決定における師管鉄沈着量の重要な役割を明らかにする ことで、NH4 + ストレスに応答する初生根 の発達の調整に寄与する関連要因について洞察を加えたものであ る。外部からのFeの利用可能性の増加は、NH4 +誘発の初生根の成長阻害を強めるという事実がある。したがって、根におけるNH4 +毒性の発生は単一要因ではなく、Feもこのプロセスに関与している。注目すべきは、NH4 +の形成に有利な土壌の条件は、Feの可溶化にも有利であるということである。例えば、無酸素状態の湿地では、可溶性Feのレベルは容易に10 mg kg-1 (~180 µmol kg-1)を超える。硝化よりもアンモニウム化の割合が高いためにNH4 +に富む酸性土壌では、低いpHもFe溶解度を大きく上昇させる。植物と土壌における NH4 + と鉄のこの密接な関係は、NH4 + ストレスに対処する農法や遺伝子操作の植物を開発する際に考慮されるべきものである。
フェロキシダーゼ(LPR2)は、NH4 +による師管への鉄の沈着に重要である。予想に反して、LPR2が中心柱に分布する領域は、師管に特異的に鉄が沈着する領域よりもはるかに広いことがわかった。LPR2は主に細胞壁マトリックスに存在するため、師管細胞特異的な鉄排出によるLPR2基質の供給源が、NH4 +による師管での鉄沈着の特異性に影響を与えている可能性が考えられる。根の細胞タイプ毎のマイクロアレイ発現プロファイルの包括的なマップは、FPN1(フェロポーチン)が主に師管細胞で発現していることを示した。したがって、Fe2+排出タンパク質であるFPN1も、Fe沈着の師管特異性を制御する役割を担っている可能性がある。それにもかかわらず、T-DNA fpn1挿入変異体においてFPN1を破壊しても、lpr2変異体のようにNH4 +ストレス下でのPRの成長は改善されず、上記のFPN1の想定される役割には反している。
もう一つの可能なメカニズムは、師管を経由して光合成産物が配分されることで、師管細胞およびその隣接細胞の細胞壁マトリックスがより多くの還元剤を蓄積し、LPR2-触媒反応のためのFe2+を生成することが有利となる可能性である。師管の細胞壁マトリックスにおける鉄の起源にかかわらず、NH4 + ストレスによる師管での鉄の大量沈着は、LPR2 の作用に密接に依存している。興味深いことに、鉄は無機リン酸(Pi)欠乏に応じた初生根の再形成にも重要な役割を果たしており、これは主にLPR1の作用に依存している。しかし、LPR1 と LPR2 のアミノ酸配列は 79% 同一であるが、LPR2 のみが NH4 + を主な窒素源とする存在下で初生根の発達を再編(阻害)する。
LPR1 の発現を LPR2 の作用部位に限定しても lpr2 変異体の NH4 + 感受性表現型が回復しないことから、植物における LPR1 と LPR2 の酵素作用は異なる制御を受けている可能性があることが示唆された。さらに、LPR1 の組織分布も LPR2 の組織分布と異なっている。これらの違いは、初生根 の NH4 + 感受性を媒介する LPR2 の特異性を説明するものであると思われる。重要なことは、リコンビナント GST-LPR2 の Km 値 (~17 µM Fe2+) が LPR1 のそれ (~3 µM Fe2+) よりも大きいということである。この相違は、Pi 欠乏や NH4 + ストレスに対する 初生根 の成長反応を制御する際の鉄の用量要件の相違とよく対応している。LPR1 が Pi 欠乏下で 初生根の成長を抑制するには 2.5 µM の鉄で十分であるが、LPR2 が NH4 + ストレス下で 初生根の成長を抑制するには 10 µM 以上の鉄が必要であった。
鉄は、NADPH オキシダーゼ、チトクローム P450 酵素、リポキシゲナーゼ、キサンチンオキシダーゼなど、いくつかの活性酸素生成酵素の補因子である。また、鉄の酸化還元サイクルは、フェントン反応を介して直接的に活性酸素を発生させることが可能である。従って、鉄の異常蓄積は、しばしば、直接的、間接的に活性酸素のバーストを引き起こす。鉄が活性酸素の発生を制御する手段が何であるかにかかわらず、我々の研究では、NH4 +による鉄の沈着の結果、根の師管で活性酸素が明らかにバーストした。しかし、このバーストが師管の細胞内または細胞外マトリックスのどちらで発生したものかを区別することはできなかった。根の師管のアポプラストへの鉄沈着を考慮すると、NH4 +による活性酸素のバーストは細胞外でのプロセスである可能性が最も高い。これまでの研究により、カロースの沈着と外部からの生物的・非生物的なシグナルに応じた活性酸素のバーストとの間に強い関連があることが明らかになっている。ROSバーストは、NH4 +ストレスに応答したカロースの沈着の誘導にも関与している。Pi 欠乏による 初生根の成長阻害は、伸長領域と 根の幹細胞間隙(?:stem cell niche SCN) におけるカロースの蓄積の亢進と関連しており、これは主に LPR1 を介した鉄沈着に依存していることが確認された。予期せぬことに、すべての処理で SCN にも鉄の沈着が見られたが、特に鉄が少ない条件では、この領域のカロースは処理間で対応するように蓄積しなかった。
カロースはβ-1,3-グルカナーゼによって分解される。バイオインフォマティクス解析により、シロイヌナズナでは約50のβ-1,3-グルカナーゼ関連遺伝子が同定されている。最近のシングルセルRNA-Seqでは、At1g66250, At2g05790, At2g01630 (PdBG2), At4g29360, At5g42100, At5g58090などのいくつかのβ-1,3-グルカナーゼ関連遺伝子が根のSCNで優先的に発現することが示された。これらのβ1,3-グルカナーゼがSCNにおけるカロースの蓄積に与える影響は、処理によって異なる可能性がある。師管は、環境に応じて植物の生長可塑性を調節する重要なメディエーターとして関与している。通常の生育条件下では、カロースは師管の篩板sieve plates とplasmodesmata領域に基底レベルで存在する。カロースが師管に大量に沈着すると、篩孔を塞ぎ、形質膜の透過性を低下させる可能性がある。実際、エスクリンを用いた我々の細管移動プローブテストは、NH4 +によるカロースの沈着が師管輸送とアンローディングを著しく阻害する直接的な証拠を示している。スクロースは子葉や成熟葉から根までの師部における主要な輸送糖であり、根の成長は地上部分から輸送されるスクロースに大きく依存している。エスクリンはスクロース輸送体依存的に師管にロードされる。したがって、NH4 +-による師管の作用阻害は、エスクリンによって示されるように、根の成長領域へのスクロース供給が、正常な細胞分裂と分化を維持するためには不十分である可能性を示唆している。この考えは、切り離された根の上部からのスクロース補給量の上昇が、NH4 + ストレス下での初生根成長を完全に回復させたことからも支持される。
これまで、NH4 + の毒性作用として、炭素供給の枯渇、ミネラルカチオン(K+)の欠乏、NH4 + 同化に伴う根の過剰なエネルギー需要、無駄な膜貫通 NH4 + サイクルに伴う高いエネルギーコストなどのいくつかの生理学的問題が提案されていた。 しかし、これらの作用は、異常な鉄沈着による師管におけるスクロース輸送の阻害と関係していると思われる。
これは、スクロースの供給不足は直接的に炭素枯渇につながるだけでなく、過剰なNH4 +同化または無駄な膜貫通NH4 +循環のいずれかのエネルギー需要を満たすことができなくなるためである。さらに、師管へのスクロースの輸送にはK+の負荷の作用が必要であり、NH4 +ストレスによるK+の欠乏はさらに師管でのスクロース輸送の阻害をもたらす可能性がある。これらのことから,Fe の沈着に伴う師管の作用阻害が,根の発達阻害と NH4 + ストレスを結びつける主要なメカニズムである可能性がある。なお、師管は糖の輸送以外にも、タンパク質、RNA、ペプチドなど数多くの物質を輸送しているが、現在のところ、機能との関連が確認されているのはごく一部に過ぎない。最近、5-メチルシトシンで修飾されたAtTCTP1 mRNAが師管を移動することで、シロイヌナズナの初生根成長をわずかに促進することが示された。また、原師管(プロトフローエム)は環境条件を伝えるCLE(Clevata)ペプチドを感知し、それに応じて根の成長を調節する結節点であることが提唱されている。これらのことから、初生根の成長を制御する師管の働きは、単にスクロースの移動にとどまらないことがわかる。したがって、NH4 +ストレス下での師管の作用の障害による初生根の成長阻害の制御に、他の物質、特にシグナル分子がどのように関与しているかは、今後さらに研究が必要であると思われる。
以上より、支配的な窒素源の変動に対応して根の発達が再形成される機構を提案した(図8)。NH4 +は、NO3 -を主な窒素源とする場合と比較して、細胞壁に局在するフェロキシダーゼLPR2の作用により、根の葉緑体への過剰な鉄の沈着を誘導する。このような異常な鉄の蓄積は、活性酸素の発生を誘発し、結果としてカロースの大量沈着を引き起こす。このため、師管の働きが損なわれ、根の成長領域へのスクロースの分配が不十分となり、根の発達が停止してしまう。今回の発見は、根の師管への鉄の集積を遺伝子操作することによって、NH4 +に富む土壌環境に対する植物の適応を改善する戦略を提供する可能性がある。
図1. NH4+培地におけるLPR2依存的な初生根の成長阻害
a Col-0とisas, lpr2-1, lpr1-1, lpr1lpr2変異体の表現型の比較
b 初生根の伸長
c Col-0, lpr2-1と補体(COM#7とCOM#15)の表現型の比較。
d 分裂細胞の数
e 伸長細胞の数
f 最初の分化細胞の長さ
g 根の成長領域(伸長領域+分裂領域)の長さ
図2. LPR2 は細胞壁フェロキシダーゼであり、LPR1 とは代替できない。
a p35S::LPR2- GFP の根の細胞における LPR2-GFP とヨウ化プロピジウムの蛍光共焦点。 b 0.8 M ソルビトールでプラズマ分解後の p35S::LPR2-GFP の根の細胞における LPR2-GFP と FM4-64 の蛍光。4 日齢の苗を 100 µM Fe を含む NH4 + 培地に移植し、苗移植の 4 日後に共焦点解析を行った。
c 組換え GST-LPR2 のフェロキシダーゼ活性。1 μg 精製 GST-LPR2 タンパク質を用いたフェロキシダーゼアッセイ。ピンクはFe2+-フェロジン複合体を示す。基質である Fe2+ は Fe(NH4)2(SO4)2-6H2O の形で 50 μM の初期濃度で加えた。
d GST-LPR2 タンパク質の Fe2+ 濃度依存性(0-300 μM)フェロキシダーゼ活性。e, f LPR1CDS と LPR1genomic を LPR2 プロモーターで組織特異的に発現させた lpr2-1 変異株の表現型。4 日齢の苗を 100 µM Fe を含む NO3 - または NH4 + 培地に移植し、苗の移植から 4 日後に解析した。2 回 (c, d) または 3 回 (a, b, e, f) 独立した実験を行ったが、同様の結果が得られ、代表的な 1 回の実験を示す。
図3 NH4+培地における鉄依存性の一次根成長阻害。
a 種々の量の鉄を添加した NO3--N または NH4+-N 培地における Col-0 苗の一次根伸長。
b NH4 +-感受性Col-0, isas, lpr2-1, および相補 系列 COM#7 の Fe 十分濃度 (100 μM) および Fe 低濃度(10 μM) 条件下での比較。
c 初発根の鉄沈着量を Perls/DAB 染色で示した。
d Fe suff NH4+ 培地で生育した Col-0 苗の根幹における鉄沈着量の拡大図。
e Fe suff NH4+培地で生育したCol-0苗の初生根の葉茎におけるPerls/DAB染色。赤と緑の矢印は、それぞれ葉茎の側壁と篩板での鉄の沈着を示す。
f Fe suff NH4+培地で生育したCol-0苗の初生根の葉茎におけるアセトン洗浄-Perls/DAB染色。苗をNO3-またはNH4+培地に移植し,移植4日後に分析した。
図 4. 根における LPR2 の分布と NH4+ に対する応答.
a 補完系統 COM#7 の初生根における pLPR2::LPR2-YFP の発現。根をヨウ化プロピジウムで対比染色し(紫色蛍光)、YFP蛍光を分析した(緑色)。
b Fe十分NH4+培地 おけるpLPR2::LPR2-YFP発現の放射状断面(上)およびクローズアップ図(下)。相対発現量はUBQ10とEF1αの発現量の幾何平均値で正規化した。
c Col-0 根でのLPR2発現. 相対発現量は、UBQ10とEF1αの発現量の幾何平均値で正規化した。
d, e Col-0の根における代表ゲルと相対タンパク質レベル。相対的なLPR2レベルは、同じサンプルからのLPR2の信号強度とアクチンの信号強度の比から推定した。Fe十分濃度
(100 µM) または Fe低濃度 (10 µM)を添加した NO3 - または NH4 + 培地に4日齢の苗を移し、苗を移植してから4日後に分析した。各実験は独立して3回繰り返し、同様の結果を得たので、代表的な実験を示す。
図5. NH4+による鉄依存的な一次根の成長阻害は、活性酸素種(ROS)のバーストと関連している。
a, b NH4 + 源における Col-0 Fe低濃度vs. Col0 Fe十分濃度 および lpr2-1 Fe十分 vs. Col-0 Fe十分 の一対の比較で、発現が増加および減少しているデファレンシャル発現遺伝子(DEGs)のVen ダイアグラムおよび Gene Ontology 濃縮解析結果。円の大きさは遺伝子番号を表し、色はPadjust の値を超幾何学的検定で計算し、FDRを用いて多重検定で調整したものを表す。
c H2DCFDA染色によるCol-0、lpr2-1、および相補性系統COM#7実生の主根における活性酸素の可視化。
d Fe十分濃度・NH4 +培地で栽培したCol-0実生の主根におけるH2DCFDA染色のクローズアップ図。
e Fe十分濃度 NH4 +培地で生育したroGFP2-Orp1苗の初生根におけるH2O2の偽色表示と葉茎マーカーのエスクリンの蛍光の共局在。
f, g ROSスカベンジャー濃度の関数としてプロットしたCol-0苗の初生根伸長の用量-反応関係。h 750 µM dimethyl thiourea (DMTU) および 200 µM 4-hydroxy-TEMPO (TEMPO) の Col-0 実生への影響を示す画像である。標記遺伝子型の4日齢の苗を、標記活性酸素消去剤を含む、または含まないFe十分濃度(100 µM)またはFe低濃度(10 µM)を加えたNO3 -またはNH4 +培地に移した。解析は実生移植から4日後に行った。実験は独立して3回繰り返し、同様の結果を得た代表データを示す。
図6. NH4+に応答した活性酸素種を介した葉面における鉄依存的カロース沈着。
a Col-0、lpr2-1および相補系COM#7実生の原根におけるアニリンブルー染色によるカロース検出。
b Fe十分濃度・ NH4 +培地で生育したCol-0苗の初生根におけるアニリンブルー染色の放射状断面(上)およびクローズアップ図(下)
c pSUC2::GFP-sporamin 根におけるGFP-sporamin とアニリンブルー染色の蛍光共焦点化
d ROS スカベンジャーによりNH4 +誘発カロース沈着が停止された。4日齢の苗をNO3 -またはNH4 +培地に移し、Fe十分濃度(100 µM)またはFe低濃度(10 µM)と標記活性酸素除去剤の併用または非併用で処理した。各実験は独立して3回繰り返し、同様の結果を得たので、代表的な実験を示す。
図7. NH4 +培地における初生根の師管作用の阻害とスクロースによる補完作用。
a, b エスクリンの葉面輸送速度と葉面アンローディング。Col-0、lpr2-1、および相補性系統 COM#7 の 4 日齢の苗を、Fe十分濃度(100 µM) または Fe低濃度 (10 µM) を含む NO3 - または NH4 +培地に移植し、苗を移植してから 4 日後に分析。
c 局所的にショ糖を供給した際の切り離した原根の成長応答。挿入図:局所的なスクロース供給に対する切り離された原根の成長反応解析を示すスキーム。4 日齢の苗から根(1 cm)を切り取った。次に、剥離した根を、Fe十分濃度または Fe低濃度を含む NO3 - または NH4 + 培地の垂直2層分割寒天系に移し替えた。上層は様々な用量のスクロースで処理し、下層はスクロースを含まないようにした。解析は苗を移植してから3日後に行った。各実験は独立して3回同様の結果を繰り返し、代表的な実験を示した。Col-0, lpr2-1, および相補性系統 COM#7 の 4 日齢の苗を、Fe十分濃度 (100 µM) または Fe低濃度(10 µM) を含む NO3 - または NH4 +培地に移植し、苗を移植してから 4 日後に分析。
図8. 根の発達の再構築と一次窒素源であるNH4 +を結びつけるメカニズムの模式図。NH4 +を主要なN源として供給すると、NO3 -を主要なN源として供給する場合と比較して、根の葉身のアポプラストに過剰な鉄の沈着が誘導されるが、その触媒反応は、根の中心柱の細胞壁に局在するフェロキシダーゼLPR2の作用によってFe2+がFe3+に酸化されることに依るものである。このような異常な鉄の蓄積は、活性酸素のバーストを引き起こすことで葉茎の輸送と荷降ろしを阻害し、その後、カロースの大量沈着を引き起こす。その結果、スクロースが根の成長部分に十分に供給されなくなり、根の発達が停止する。
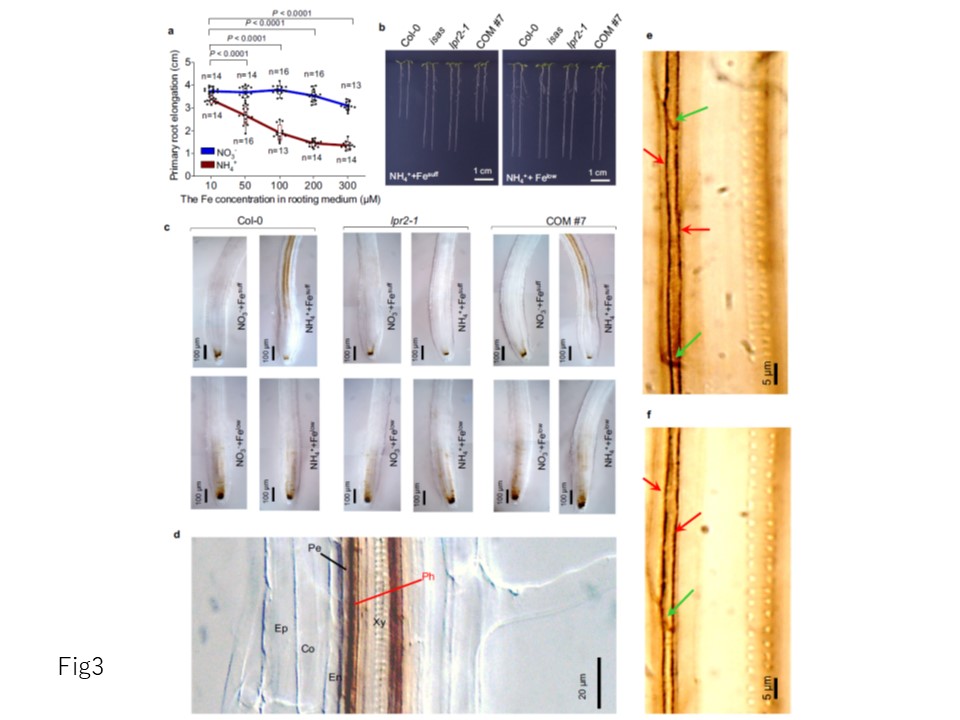
図3
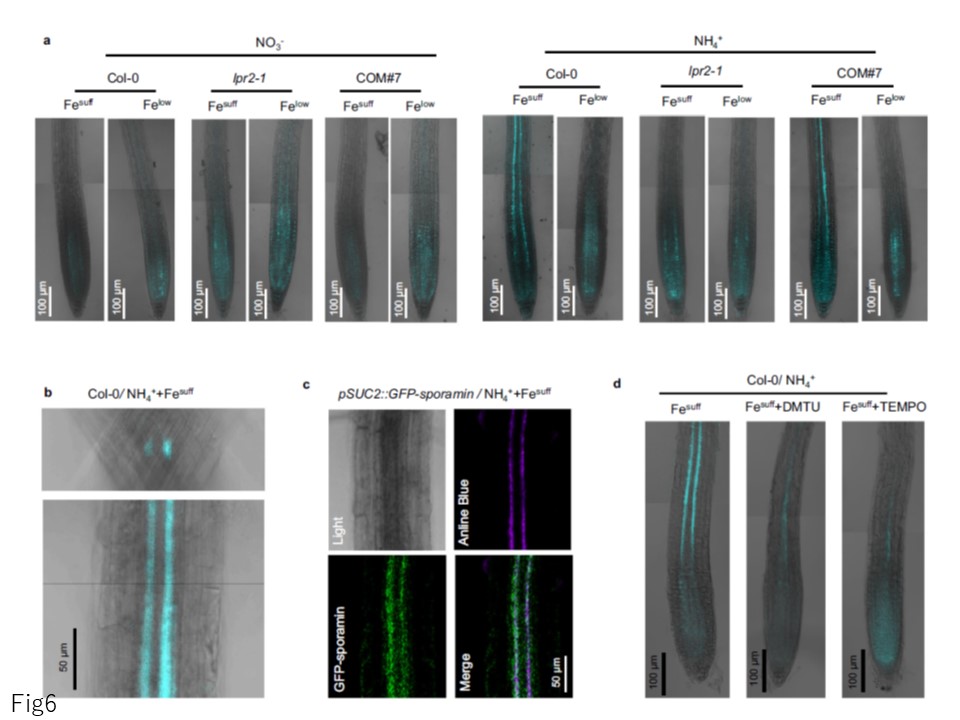
図6
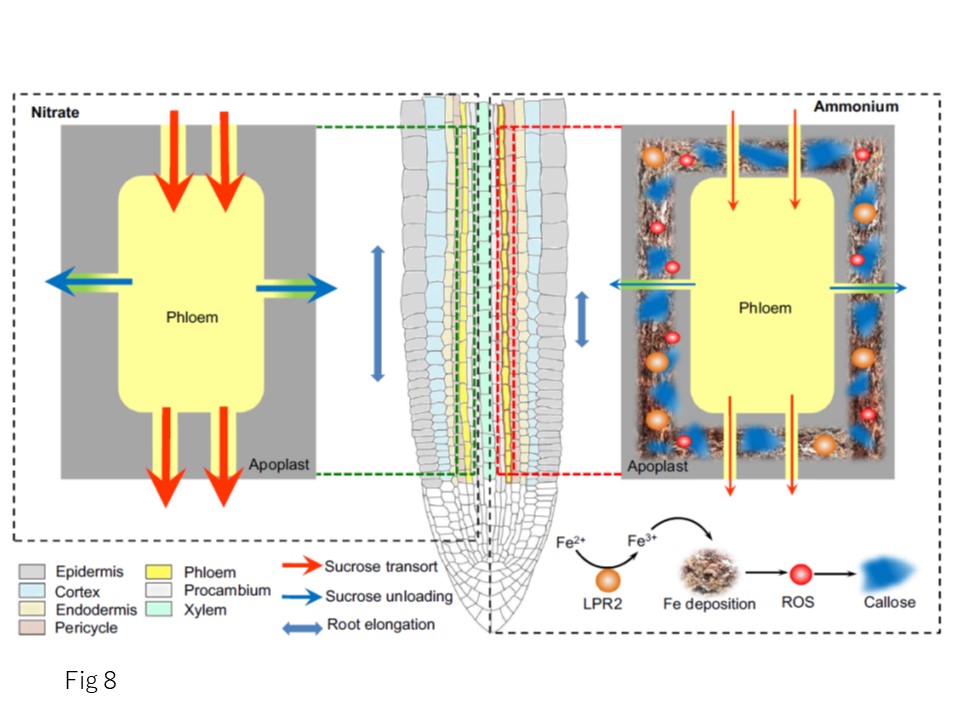
図8