microRNA408を操作して発現させるとアラビドプシスの鉄欠乏応答に影響が出る
以下の論文は英語の構文が不正確で、内容もかなり難解であったので、訳出に非常に苦労した。しかしマイクロRNAと植物の鉄欠乏の関係を述べているのが、小生には新鮮であった。読者の理解を深めるために、<要旨>だけでなく、<緒言>も全訳しておいた。
(タイトル)
microRNA408を操作して発現させるとアラビドプシスの鉄欠乏応答に影響が出る
The Altered Expression of microRNA408 Influences the Arabidopsis Response to Iron Deficiency
Àngela Carrió-Seguí, Omar Ruiz-Rivero, Laura Villamayor-Belinchón, Sergi Puig, Ana Perea-García, and Lola Peñarrubia
Front Plant Sci. 2019; 10: 324.
Published online 2019 Apr 2. doi: 10.3389/fpls.2019.00324
(要旨)
MicroRNAsは植物の全身性金属栄養ホメオスタシスに影響を与えることによって植物が様々な環境に適応することに貢献している。
銅と鉄の欠乏は相反的にアラビドプシス(Arabidopsis thaliana)のmicroRNA408 (miR408)の発現をコントロールする。
このmiR408はlaccase-like multicopper oxidase family members であるLAC3, LAC12, LAC13の転写後制御をおこなうものである。
本研究ではmiR408 T-DNA 挿入変異株 (408-KO1 と 408-KO2)と、あらかじめ特性を同定していたmiR408 の過剰発現株(35S:408-14)を用いて、miR408が銅と鉄による代謝にどのように影響しているのかを探索した。
miR408を操作した発現によって、植物のパフォーマンスや鉄欠乏条件下での鉄制御遺伝子群の活性が減少させられた。
miR408が標的とするラッカーゼの低い発現と呼応して35S:408-14植物では維管束梢のリグニン化が低下した。
鉄欠乏条件下では野生株ではphenoloxidase と ferroxidase活性の低下が起きるのだが、408-KO1(ノックアウト)植物ではラッカーゼが強く発現したためか、それが起きなかった
最後に、408-KO1株と35S:408-14 株では、鉄欠乏条件下で過酸化水素レベルが変化するのを観察した。
以上を総合的に考えると、鉄欠乏条件下でmiR408のレベルをいろいろ変化させると、多様な制御不全が起こるということが示唆された。
(緒言)
様々な栽培条件下にある植物では金属栄養のホメオスタシスを維持するために、根部と地上部間の全身性シグナルが必要である。
植物全体の適応のためには栄養素そのものばかりでなく、そのほかの分子、例えばmicroRNAs、伝達物質類、シグナル類など栄養状態の情報が必要である。
アラビドプシスではCu-miRNAsと呼ばれる一群のsmall RNAsが保存されており、銅欠乏条件下でアップレギュレートされ、必須ではない銅タンパクを標的としている。
Cu-miRNAs (miR397, miR398, miR408)は 高く保存されており、これらはsmall RNA-が媒介する植物の銅ホメオスタシス制御に重要である。
局所的な銅欠乏応答の制御とは別に、Cu-miRNAsは師管経由の全身性シグナルとして働き、銅の配分と金属欠乏に適応している。
miRNA発現における拮抗的制御を含む、異なるプロセス間での、銅と鉄のホメオスタシスには相互作用があることが実験的に示されている。
miR398はCSD1 と CSD2のmRNAsを制御するが、これらはそれぞれ細胞質とクロロプラストのCu/Zn superoxide dismutasesの遺伝子をコードしている。
銅欠乏条件下では転写因子であるSQUAMOSA-PROMOTER BINDING-LIKE PROTEIN 7 (SPL7)がmiR398をアップレギュレートするが、これはCu/ZnSOD のカウンターパートであるFe superoxide dismutase (FeSOD)と交代して、例えば銅の必須タンパクであるプラストシアニンなどに必要な銅を節約するためだと思われる。
逆に、鉄欠乏条件下ではmiR398のダウンレギュレーションはFeSODをCu/ZnSODと交代させるのに貢献する。
他に、師管液中での銅と鉄の欠乏により拮抗的に制御されるmiRNAにはmiR408がある。しかしその生理学的意味は未解明である。
アラビドプシスでのmiR408の発現は豊富であり場所が遍在している。
miR408は適切な栄養成長に必要であり、さまざまな非生物的ストレスへの適応に関係している。
銅欠乏条件下でのmiR408 による制御はmiR408のプロモーター内のGTAC モチーフと結合するSPL7によって媒介される。さらにmiR408は、光に対する応答を媒介する転写因子であるHY5 (elongated Hypocotyl 5)の標的でもある。
miR408の過剰発現は、低銅条件下でのspl7 とhy5 の変異株の効果をプラストシアニン作用を向上させることによって部分的に補償する。
HY5-SPL7 遺伝子ネットワークを通して光や銅に応答するという役割に基づいてmiR408を考えると、miR408は、光合成のために必要なプラストシアニンに適切な銅を供給する環境シグナルの集積者として働く、という役割が提唱されている。
であるから、構成的なmiR408の発現は、広域にわたる植物種の光合成のパフォーマンスを向上させ、クロロプラストの銅含量を増やし、バイオマスと種子の収量を向上させるのである。
Cupredoxin, plantacyanin, uclacyanin などのmRNAsはmiR408の標的である。これらは青色銅タンパクとしてタンパク質間の電子伝達シャトルとして働いている。
miR408はlaccase-like multicopper oxidases (LMCOs)をコードしているLAC3, LAC12, LAC13遺伝子のmRNAを標的としている。
LMCOsは細胞外glycoproteinsで、in vitroでは多くの基質を酸化触媒する、それと同調して酸素を還元する。
基質が広域にわたるので高等植物のLMCOsは、当初予想されていた作用よりもより多様な働きをし得るのではないかと提唱されている。
ある種のLMCOの作用は、リグニン化、フェロキシダーゼ活性、酸化ストレス、などの鉄ホメオスタシスに影響する過程に関係しているかもしれない。このことがmiR408にる鉄欠乏条件下でのLMCOの制御を説明するのかもしれない。
ある種のLMCOsは、二次的細胞壁形成において、リグニンモノマーからの酸化的高分子リグニン化に関係している。そしてその活性はmiRNAの発現によって影響を受ける。
cross-linkingを通しての細胞壁の修飾は金属のキレート化とその移動性に影響を与えることが示唆されている。
銅と鉄の間のホメオスタシスの間には潜在的な交互作用があり、われわれは飢餓条件下では長距離輸送の間にリガンド(配位子)に対して金属間での拮抗が起こることを見出している。
Laccasesに加うるにH2O2を基質とするperoxidasesもin vitroではリグニン生合成に貢献しており、H2O2そのものが細胞壁の架橋や弛緩に役割を果たしている。
培養液からH2O2を取り除くとドイツトウヒで形成される細胞外リグニン含量が有意に減少し、活性酸素(O-2)の合成を阻害するか、またはsuperoxide dismutasesによる活性酸素のH2O2へのdismutationはリグニン含量を減少させる。
イネのOsLAC3はH2O2の集積を誘起し、種子のsetting rate、維管束組織や根の先端のミトコンドリアの統合性に影響する。
OsLAC3 と L-ascorbate oxidasesが驚くほど似ていてもH2O2除去を抑制するアスコルビン酸酸化の役割は可視的表現型の違いで説明されると提案されている。
活性酸素種(ROS)も金属欠乏条件下での銅と鉄の相互作用に影響する。
金属間のストレスがあるときのROS、特にH2O2、の生成と消去というデリケートなバランスは拡散するシグナル伝達経路にとって重要であり、H2O2と植物の金属耐性には相関があるという証拠がある。
ある種のLMCOsがferroxidase活性を有すると推定されているが、その役割は、もう一つの銅と鉄の間のホメオスタシスの交互作用に関与しうるmiR480制御過程が存在するということであろう。
LMCOsは植物以外の生物で鉄の輸送に関与している。
ある種のLMCOは鉄の稼働化と輸送に必須な酸化還元サイクルに関与している可能性があるが、それはそれら(のタンパクが)がこの目的のための分子残基を有しているからである。
いろいろな因子のなかでも、鉄の酸化還元状態はferroxidase と ferrireductaseの比に依存している。
しかし、LMCOsが植物でferroxidasesとして働いて金属酸化に関わっているという、実験的証拠は少ない。
イネ科以外の多くの植物は細胞膜ferric chelate reductaseでFe3+ chelatesから還元して鉄を獲得している。この酵素はアラビドプシスではFERRIC REDUCTASE 2 (FRO2)にコードされている。還元の後にFe2+はIRON REGULATED TRANSPORTER 1 (IRT1)としてコードされている輸送体を通して細胞内に取り込まれる。
IRT1の基質利用性は無機や有機の鉄複合体ではないフリーの細胞外の鉄と、Fe2+/Fe3+の比に依存している。これらは細胞外の酸化還元状態とferrireductases酵素活性に依っている。
Helix-loop-helixタイプの転写因子FIT (bHLH29)は鉄獲得と可動化に関係している。
他のbHLH サブグループである Ib factors (bHLH38, bHLH39, bHLH100, and bHLH101)はFITと協奏して働きうる。
本研究ではmiR408レベルを変化させてみて、鉄欠乏が幼植物に与える影響を、LMCOのmRNAレベル、酵素活性、鉄欠乏応答性を分析することによって探索した。
(下図の説明)
銅と鉄のホメオスタシスがmiR408を通して交互作用するモデル。本実験において観察された鉄欠乏応答に対する主なmiR408効果の概念図
miR408は直接の標的遺伝子であるLMCOs、 LAC3, LAC12, LAC13 (LAC3/12/13)、 cyanins (i.e., ARPN) のmRNAの発現に影響を与え、これらのリグニン合成、phenoloxidase, ferroxidase, ascorbate oxidaseなどの酵素活性に対する様々な役割に影響する。
他の非生物ストレスに加えて、銅欠乏と鉄欠乏はmiR408による制御とその後の酵素活性に正反対に作用する。
鉄欠乏条件下でのFIT依存性やFIT非依存性のシグナル経路はmiR408によって前者で誘導されたり、後者で抑制されたりする。
鉄欠乏誘導性酸化ストレスと過酸化水素(H2O2)は可視的なmiR408効果の中で役割を果たすことができる。
H2O2はLMCO酵素活性に影響し、少なくともmiR408作用の一部は鉄欠乏下でのこの効果を打ち消すことができうる。
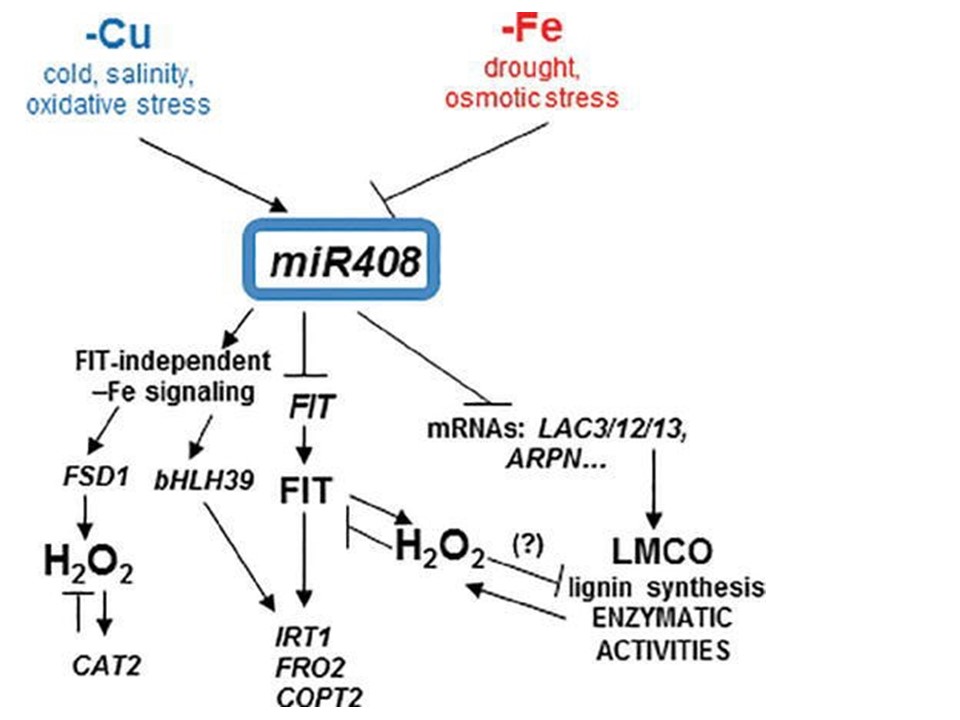
銅と鉄のホメオスタシスがmiR408を通して交互作用するモデル