コメント:植物の鉄ホメオスタシスの負と正を連結するメカニズムについて
植物の鉄ホメオスタシスの負と正を連結するメカニズム
Connecting the negatives and positives of plant iron
homeostasis
New Phytologist (2019) 223: 1052–1055
www.newphytologist
以下の図1の説明.ILR3は鉄(Fe)ホメオスタシスの負の制御因子であるとともに正の制御因子としても作用する
低濃度鉄条件下では(左)ILR3は強力に誘導されたPYEと二量体を形成してフェリチン遺伝子の発現を抑制し、細胞内の利用可能な大部分の鉄を、鉄要求性タンパクに提供する。
同時にILR3はbHLH クラスの IVc タンパク (bHLH IVc)と共に土壌からの鉄吸収機構を正に制御する。
鉄要求性タンパクに対して鉄が十分に利用できる鉄過剰条件下では、bHLH クラス IVc タンパク (bHLH IVc)は抑制されて、フェリチンの発現が増加し過剰のフリーの鉄を隔離する。
同時に、根からの鉄吸収活性は低くなる。このことによって植物がフリーの鉄によって触媒される活性酸素種(ROS)の生成をコントロールする手助けをしている。
以下の図2の説明.全身的な鉄(Fe)ホメオスタシスの全体像
根からの鉄吸収の正の制御因子は緑色に、負の制御因子は赤で示している。
細胞内でのフリーの鉄は鉄吸収の負の制御因子である。鉄欠乏に応答するときは植物は鉄吸収機関(IRTコホート)を根の表皮細胞でup-regulateする。
このIRTコホートは4つの膜タンパクを意味している:根圏を酸性化するAHA2、鉄と結合するクマリンを排出するPDR9、鉄を還元するFRO2、Fe(II)イオンを吸収するIRT1である。
鉄吸収遺伝子の発現を制御しているのは、より下位のbHLH38/39と結合しているFITタンパクである。
より上位のbHLH34, bHLH104, bHLH115 , ILR3(すべてIVc bHLHタンパク分枝に属する)の二量体が下位のbHLH38/39の遺伝子発現を誘導する。
このカスケードはBTSL ユビキチンライゲースによって負に制御されている。
低濃度鉄条件下では上位のbHLHタンパク(bHLH34, bHLH104, bHLH115 , ILR3)がpericycleでも同様に発現して、bHLH38/39 と bHLH100/101の発現誘導している。これらの転写因子の直接の標的については本文で広範囲述べておいた。さらに、PYE と ILR3は複合体を形成し局所的な鉄のプールを低下させるプロセスをdown-regulateする。
PYE/ILR3による転写レベルでの負の制御の標的は3種類のフェリチン(FER)、NAS4ニコチアナミン合成酵素(これは維管束への鉄の附加を容易にする)、およびその他の鉄の局在を媒介する膜結合顆粒タンパクである.
BTS1も内鞘での負の制御因子である。葉においてはOPT3は導管から師管への鉄の移行を媒介する。
YSL1/3は師管から葉の柔組織のアポプラストへ鉄を排出し、そこから光合成に多量の鉄を要求する細胞に鉄が入っていくのだろう。
OPT3が根からの鉄吸収の負の制御因子であり、YSL1/3が鉄吸収作用の正の制御因子であるというこのモデルは、これまでの文献と合致するものであるが、これらの輸送体作用のメカニズムはまだ十分には解明されていないことに留意されたい。
光合成は鉄によって強く制限されており、また環境因子によっても巧妙に制御されているので、クロロプラストの鉄・タンパクの高い発現が鉄吸収の正の制御因子たりえて、他の細胞内鉄含有タンパクはむしろより構成的な作用をしているのだろう。
鉄欠乏下での師管での鉄レベルの調節を考える場合、YSL1/3 と OPT3では、前者がdown-regurateされており、後者がup-regulateされている。
鉄欠乏では師管での短いペプチド類、FEP/IMA、の発現が増加し、これらは鉄吸収に対しては正の制御因子である。
鉄欠乏下では、鉄はまずミトコンドリアの活性が優先し、プラスチドのFERの発現がdown-regulateされ、光合成タンパクに利用能性の鉄が増加し、その他のクロロプラストで鉄要求性のタンパクに必要な鉄が増加する。
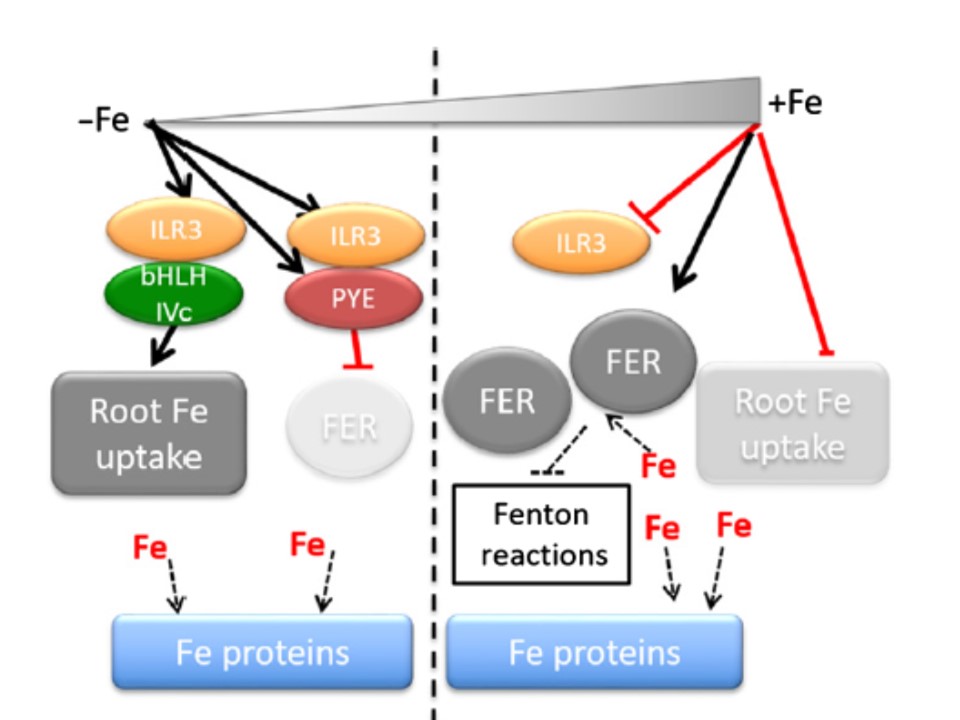
図1
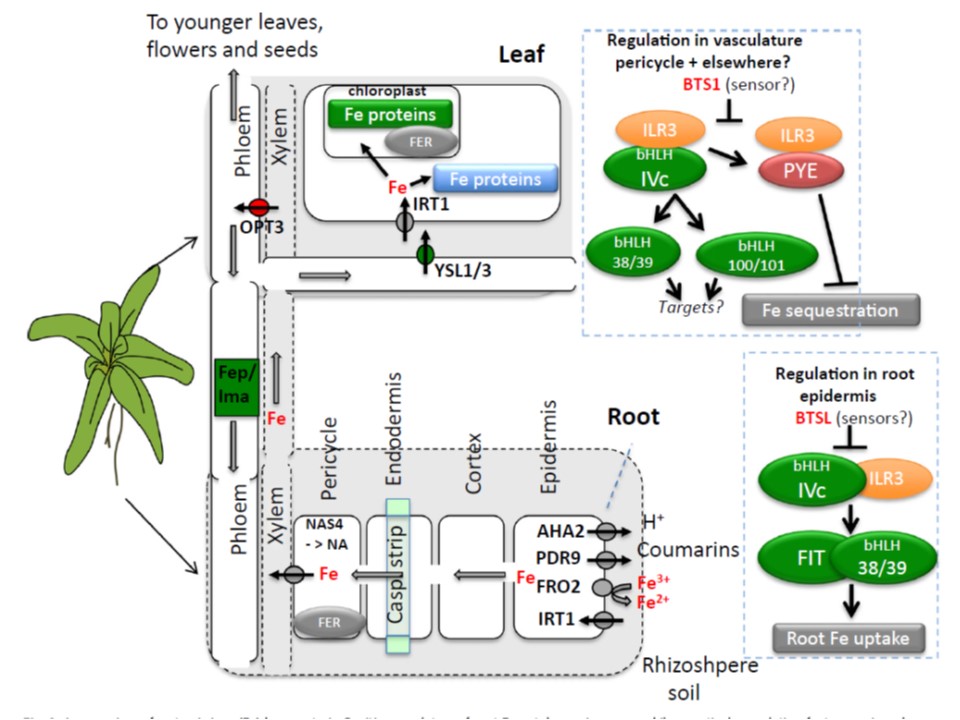
図2