鉄と活性酸素種に依存した「イネーイモチ菌」相互作用による Ferroptotic Cell Death(フェロトーシス)
以下の論文は、鉄と病原菌感染症との関係について詳しく述べたものである。要約 ばかりでなく 重要だと思ったので まえがき も訳してみた。著者らのくどくどしい英語表現のまずさもあって、なかなか翻訳しにくかったのだが、大略理解してもらえると思う。
(タイトル)
鉄と活性酸素種に依存した「イネーイモチ菌」相互作用による Ferroptotic Cell Death(フェロトーシス)
Iron- and Reactive Oxygen Species-Dependent Ferroptotic Cell Death in Rice–
Magnaporthe oryzae Interactions
Sarmina Dangola, Yafei Chena, Byung Kook Hwangb, and Nam-Soo Jwaa,*
Plant Cell Advance Publication. Published on December 18, 2018, doi:10.1105/tpc.18.00535
©2018
(要約)
超感受性応答(HR)細胞死は、カビの寄生的侵入を抑制する最も効果的な植物の免疫応答である。
この論文で我々はイネ(Oryza sativa)とイモチ病菌(Magnaporthe oryzae)の相入れない相互作用において、イネの細胞が鉄と活性酸素種(ROS)依存性のフェロトーシスによる細胞死が誘導されることを報告する。
非発病性のイモチ病菌に感染しているときにはHR細胞死が進行しているイネの葉鞘組織に鉄イオンとROS(例えば過酸化水素)が蓄積した。
それに対して、発病性イモチ病菌に感染しているときや、カビのエリシターであるキチンで処理した場合には鉄はイネの細胞に集積しなかった。
ΔOs-nadp-me 2-3変異株に対して病原性イモチ病菌を感染させても鉄やROSの集積の引き金が引かれず、HR細胞死を抑制した。このことは、イネにおいてはNADP-malic enzyme 2がフェロトーシス細胞死に必要であることを示唆している。
イネの葉鞘において病原性イモチ病菌に感染しているときに小分子のフェロトーシス阻害剤であるdeferoxamine、 ferrostatin-1および cytochalasin、また NADPH oxidase 阻害剤である diphenyleneiodonium は、鉄依存性ROS集積と脂質過酸化を抑制し、HR細胞死に至るのを完全に軽減した。
これに対して、小分子である誘起材 erastin は、鉄依存性ROS集積とグルタチオン欠乏の引き金を引き、それが結果的にはイネにおける発病性イモチ病菌によるHR細胞死に導くことになった。
これらを総合すると、イネにおいてイモチ病菌による感染を中断させるフェロトシス細胞死の過程には、鉄とROSに依存性のシグナルカスケードが関係している、ということを示している。
(まえがき)
侵入する微生物病原体に対して植物が細胞死を遂げることは効果的な免疫防御反応として決定的である。
宿主植物は病原体の攻撃に対して細胞死を引き起こすことができるが、それは侵入部位における病原体の成長と増殖を抑制する作用を行う。
スーパーオキサイド (·O2 −), 過酸化水素(H2O2), ハイドロキシラジカル (·OH)などの活性酸素種(ROS)は、細胞死と免疫力達成への誘導、シグナル化に関わっている。
ROSの爆発は植物細胞が病原体を認識する最も早期に起こる防御シグナルである。
ROSは当初はアポプラストで生産され、病原体に対する抵抗反応を促進するために直接細胞壁を強化する。
弱い一時的なROSの爆発は病気を起こす有毒な(親和性の)病原体との相互作用の間に起こっている;しかし、強い持続性のROS爆発は非病原性の(非親和性の)病原体によって誘導され、抵抗性と過敏感応答(HR)による細胞死を引き起こす。
病原体関連分子パターン(PAMPs)を認識する、いくつかのパターン認識レセプター(PRRs)が植物細胞膜に同定されてきた。
非親和的な植物・病原体相互作用では、抵抗性宿主の細胞内でのヌクレオチド結合性ロイシンリッチリピート(NLR)リセプター が特異的病原体のエフェクターを認識して植物におけるROS爆発と急激なHR細胞死に至らしめる。
フェロトーシスとは、制御された非アポトーシス型の鉄依存性の細胞死で、最近動物細胞で発見されたものである。
フェロトーシス的細胞死はアポトーシス、ネクローシス、オートファジーなどとははっきりと異なるものである。
フェロートーシスはグルタチオン依存性酸化防止のメカニズムを不活性化し、その後の鉄依存性毒性脂質ROSとりわけ脂質過酸化物(lipid hydroperoxides)の集積によって引き起こされる。
ROS、鉄、脂質過酸化物は直接的にフェロトーシスの過程に関与している。
人間と病原性微生物において、感染や免疫における多様な細胞プロセスにおいて、鉄は酸化還元の触媒として働き、電子の授受を行う。
植物の根の伸長過程では、細胞特異的なアポプラストの鉄とかロース沈積が起こり根の生長点の維持を調整しているが、これは細胞と細胞のシンプラストの情報交換(対話)にも似ている。
最近の研究では植物において熱(ヒート)ストレスがフェロトーシス様細胞死を誘導したと報告された。
非親和性の植物―病原体関係では急速なROS、鉄、α-glutamylcysteine合成酵素の増加現象がフェロトーシスの重要なマーカーであるといってもよいかもしれない。
低分子のフェロトーシス阻害剤であるdeferoxamine (DFO) や ferrostatin-1 (Fer-1)は哺乳類のフェロトーシス経路におけるROS依存性細胞死を抑制する。
DFOはバクテリアの鉄キレーターで、効果的に鉄を細胞内に収着してフェロトーシス細胞死を阻止する。
フェロトーシス阻害剤であるFer-1は、鉄依存性ROS集積による脂質過酸化を阻害する。
フェロトーシスは腫瘍性RAS(HRASG12V)変異細胞株を選択的に殺す作用がある低分子エラスチンによって誘導される。
エラスチンは細胞膜上のcystine/glutamate antiporter (system Xc−)活性を得意的に阻害する。それにより細胞内へのcystine吸収をかく乱して細胞内グルタチオン(glutathione)を欠乏させ、glutathione peroxidase 4 (GPX4)を不活化するのである。
グルタチオンは強力なアンチオキシダントである;すなわちグルタチオン欠乏は細胞内ホメオスタシスをかく乱し、ROSの集積を起こすのである。
増強されたROSは細胞内鉄と反応して毒性の強い脂質過酸化物(lipid peroxides)を形成するのである。
In vitro 実験でも、エラスチンは細胞のROSレベルを異常に亢進し、鉄とROS依存的な過剰な脂質過酸化によってフェロトーシス的細胞死を誘導する。
非親和的なイネ(Oryza sativa)-イモチ病菌(Magnaporthe oryzae)の交差反応は、植物における病原誘導性の高感受性(HR)細胞死や防御反応の根底にある分子レベルや生化学レベルの機構を同定する好都合の実験系である。
非病原性のM. oryzaeによる感染に応答して、イネにおいてアポプラストのROS爆発が起こり高感受性(HR)細胞死を誘発する。
植物細胞では、スーパーオキサイドデイスムターゼ (SOD), カタラーゼ (CAT),アスコルビン酸パーオキシダーゼ
(APX), グルタチオン還元酵素(GR),グルタチオン S-トランスフェラーゼ (GST),グルタチオンパーオキシダーゼ (GPX)らがROS (·O2 −と H2O2)の無毒化に重要な役割を担っている。
細胞質NADP-malic enzyme (ME)はNADPH酸化酵素に電子(e-)を供与するアポプラストのROS生成に必要で、このROSが最終的にイネの高感受性(HR)細胞死に至らしめるのである。
NADPH酸化酵素の阻害剤であるdiphenyleneiodonium (DPI)は、哺乳類細胞のフェロトーシス細胞死を抑える。なのでNADPH酸化酵素は細胞死誘導型ROSの供給源である。
イネ細胞のキチンエリシター結合型タンパク(OsCEBiP)は、カビの細胞壁エリシターキチンを、キチンエリシターリセプターカイネース1(OsCERK1)とともにヘテロオリゴマーリセプター複合体の形で認識し、これがROS爆発の引き金を引くのである。
イネのOsCERK1遺伝子破壊株は完璧にキチンが引き金となるROS (H2O2)の生成を破壊するので、イネ葉鞘細胞での非病原性のイモチ病菌M. oryzaeの菌糸の成長に導くのである。
M. oryzaeのエフェクターであるAvrPiiはNADP-ME活性を直接阻害することによってイネのROS生成を制限する。
この研究で我々は、鉄とROS依存性シグナルカスケードが、非親和性イネーイモチ病菌M. oryzae相互作用におけるHR細胞死に関わっていることについて報告する。
鉄はフェロトーシス細胞死を引き起こす脂質過酸化物の集積に必須である。
無毒性イモチ病菌M. oryzaeに感染したイネ葉鞘表皮細胞では鉄(Fe3+)とROS(H2O2)が細胞内と侵入菌糸の周りに集中的に集積する;しかし毒性菌糸に感染したイネ細胞には集積しない。
我々はここでNADP-malic enzyme 2(NADP-ME2)の存在が、鉄-、ROS-依存性フェロトーシス細胞死に決定的であることを示す。
デフェロオキザミン(deferoxamine)、フェロスタチン( ferrostatin-1), サイトカラシンE(cytochalasin E), NADPH オキシダーゼ阻害剤 DPIなどの低分子のフェロトーシス阻害剤, そしてRAS選択的致死性低分子誘起剤エラスチン(erastin)を用いて、鉄依存性ROS集積と脂質過酸化が、無毒性イモチ菌M. oryzae によるHR(過敏感応答)細胞死に関係しているかを調べた。
毒性イモチ病菌M. oryzaeにおいて、エラスチン処理により鉄とROSの集積が見られたが、グルタチオン生産が阻害され、最終的に鉄-ROS-依存性フェロトーシス細胞死となった。これは無毒性イモチ病菌M. oryzaeによって誘導されるHR(過敏感応答)細胞死と同様であった。
イネ細胞においては、カビの細胞壁エリシターであるキチンはROS爆発の引き金を引くが、鉄の集積は行われず、HR(過敏感応答)細胞死にはならなかった。
結果を集約すると、ROS-依存性フェロトーシスがイネとイモチ病菌のHR(過敏感応答)細胞死に関係していることを示唆している。
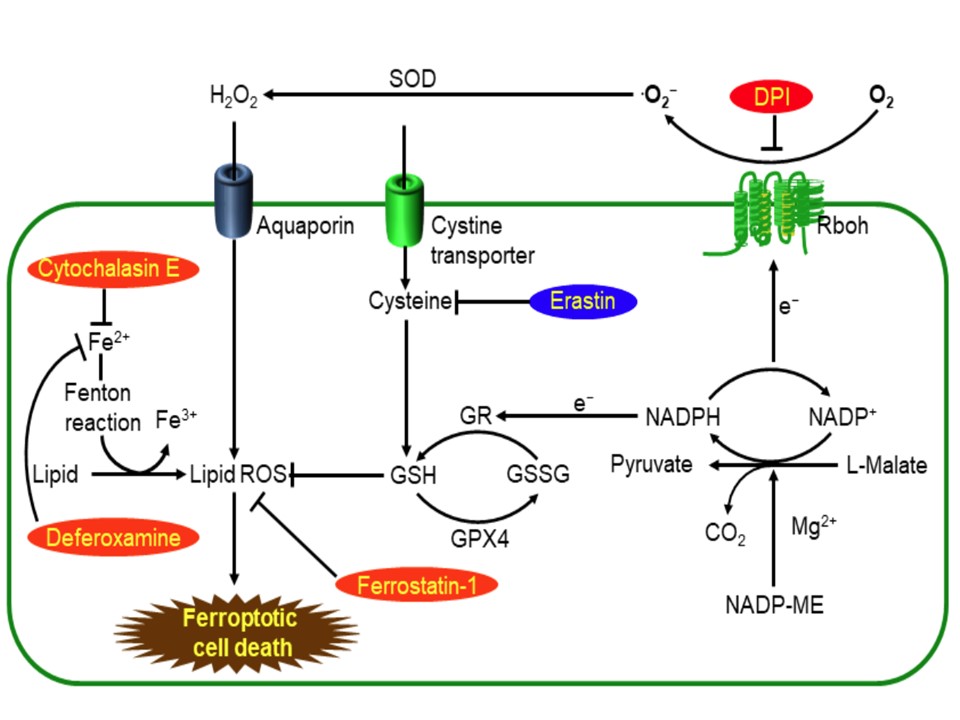
(下図の説明)
イネ-イモチ病菌相互作用におけるROS依存性フェロトーシス的細胞死のモデル
NADP-MEはNADPHをNADPH 酸化酵素 (Rboh)への
電子供与体として提供し、RbohsはROSの強固な生成に必要である。アクアポリンチャンネルはH2O2を生態膜通過させる。細胞内の高い活性鉄(Fe2+)はH2O2と反応してFe3+とハイドロキシラジカル(∙OH)を生成する。低分子阻害剤deferoxamine, cytochalasin E, ferrostatin-1, and DPIらは赤で表記している。低分子誘起剤エラスチン(erastin)は青で表記している。
エラスチンはcystine antiporter(system Xc –)を阻害してグルタチオン欠乏を誘起し、おそらくGPX4を不活化し、過剰な脂質過酸化を引き起こし、最終的に鉄-,ROS-依存性フェロートーシス的細胞死を引き起こす。
DPI, diphenyleneiodonium; GR, glutathione reductase; GSH, reduced glutathione; GSSG, glutathione disulfide, oxidized glutathione; GPX4, glutathione peroxidase 4; NADP-ME, NADP-malic enzyme; Rboh, respiratory
burst oxidase homolog; SOD, superoxide dismutase.