揝寢崌E3儔僀僎乕僗偼倐HLH揮幨場巕傪僞乕僎僢僩偵偟偰怉暔撪偺揝偵墳摎偡傞
揝寢崌E3儔僀僎乕僗偼倐HLH揮幨場巕傪僞乕僎僢僩偵偟偰怉暔撪偺揝偵墳摎偡傞
Iron-Binding E3 Ligase Mediates Iron Response in
Plants by Targeting Basic Helix-Loop-Helix
Transcription Factors
Devarshi Selote, Rozalynne Samira, Anna Matthiadis, Jeffrey W. Gillikin, and Terri A. Long
Plant Physiology_, January 2015, Vol. 167, pp. 273–286
乮梫巪乯
怉暔傗摦暔偵偍偄偰揝偺媧廂偲戙幱偼嬞枾偵惂屼偝傟偰偄傞丅
傾儔價僪僾僔僗偱偼BRUTUS(BTS)偼3偮偺hemerythrin (HHE) 僪儊僀儞偲Really Interesting New Gene (RING) 僪儊僀儞傪桳偟偰偍傝丄basic helix-loop-helix乮倐HLH乯揮幨場巕偲憡屳嶌梡偡傞揝寚朢墳摎惈偺惓偺惂屼場巕偱偁傞 POPEYE (PYE)偲僿僥儘僟僀儅乕傪宍惉偡傞丅BTS偼E3儕僈乕僛擻傪桳偟偰偍傝丄揝寚朢偵懳偡傞崻偺惉挿丄崻寳偺巁惈壔丄揝娨尦妶惈偵偨偄偡傞栶妱傪墘偠偰偄傞偙偲偑傢偐偭偰偄傞丅偙偺僞儞僷僋偺嶌梡傪偝傜偵柧傜偐偵偡傞偨傔偵ProBTS::b-GLUCURONIDASE慻傒懼偊懱傪嶌惉偟偰丄BTS偑惗怋婜偺惉挿傗敪惗偲尒偐偗忋楢摦偟偰丄敪惗拞偺泱夎傗傎偐偺惗怋慻怐偱丄敪尰偟偰偄傞偙偲傪尒弌偟偨丅
傑偨丄BTS偲PYE-like (PYEL)側bHLH揮幨場巕偼妀撪偱憡屳嶌梡偟偰偄偰丄偦傟偑BTS偺RING僪儊僀儞偵埶懚偟偰偄傞偲峫偊傜傟偨丅
傑偨丄BTS偑揝寚朢忦審壓偱PYEL僞儞僷僋偑26僾儘僥傾僝乕儉宱桼偱暘夝偝傟傞偙偲傪懀恑偡傞偲偄偆徹嫆傪帵偟偨丅
傑偨丄揝偑BTS偺HHE僪儊僀儞偲寢崌偡傞偲晄埨掕壔偡傞偑丄偦偺晄埨掕壔偼HHE僪儊僀儞撪偺摿堎揑側傾儈僲巁巆婎偵埶懚偡傞偙偲傕柧傜偐偵偟偨丅
杮尋媶偼丄揝寚朢墳摎偵娭偡傞堚揱巕敪尰惂屼僱僢僩儚乕僋偺峔惉惉暘傪E3儐價僉僠儞儕僈乕僛偱東栿屻惂屼偡傞偲偄偆怉暔揝儂儊僆僗僞僔僗偵娭偟偰廳梫偱儐僯乕僋側儊僇僯僘儉傪柧傜偐偵偟偨傕偺偱偁傞丅
乮傑偊偑偒乯
丂
揝偼懡偔偺戙幱夁掱偵娭學偟偰偄傞杮幙揑側惉暘側偺偱惗暔偵偲偭偰揝寚朢偼廳梫側栤戣偱偁傞丅摨條偵揝夁忚傕揝偑巁慺偲寢傃偮偄偰忈奞揑側妶惈巁慺庬傪惗惉偡傞偺偱栤戣偱偁傞丅偩偐傜怉暔傗摦暔偼揝偺懱撪棙梡偵懳偡傞墳摎傪尩枾偵惂屼偟偰偄傞丅
丂怉暔偼搚忞偐傜揝傪媧廂偡傞偨傔偵2庬椶偺婎杮揑側曽朄傪恑壔揑偵妉摼偟偨丅
掅揝塰梴忦審壓偱偼僀僱傗僩僂儌儘僐僔側偳偺扨巕梩(grass)怉暔偼傑偢Strategy-II偺墳摎傪偡傞丅偦偙偱偼崻寳偵
phytosiderophore 乮僼傽僀僩僔僨儘僼僅傾乯傪暘斿偟偙傟偑Fe3+偲崅恊榓揑偵寢崌偡傞丅偙偺揝丒僉儗乕僩偼嵶朎枌偵嬊嵼偡傞yellow stripe 傗yellow stripe -like(YSL)桝憲懱傪宱桼偟偰崻偵庢傝崬傑傟傞(Curie et al., 2001; Inoue et al., 2009)丅 傑偨丄偄偔偮偐偺尋媶偱偼僀僱偼傑偨捈愙Fe2+傪捈愙庢傝崬傓偙偲偑偱偒傞丅
憃巕梩怉暔偲nongraminaceous扨巕梩怉暔偼掅揝塰梴忦審壓偱偼Strategy-I偺曽幃傪棙梡偡傞丅傾儔價僪僾僔僗(Arabidopsis thaliana)偼Strategy-I偺墳摎傪帵偟悈慺僀僆儞傪捠偡H+ATPase, 傛偔抦傜傟偨AHA2偺堚揱巕敪尰偲峺慺妶惈傪憹壛偝偣偰丄崻寳傪巁惈壔偟偰崻抂嵶朎晅嬤偺巁壔揝傪壜梟壔偡傞(Römheld et al., 1984; Santi and Schmidt,2009)丅傑偨摨帪偵枌寢崌揝娨尦峺慺FERRIC
REDUCTASE OXIDASE2 (FRO2)妶惈傪崅傔偰Fe3+傪Fe2+偵娨尦偡傞(Robinson et al., 1999)丅Fe2+偼嵶朎枌偵嬊嵼偡傞IRONREGULATED丂TRANSPORTER1 (IRT1)偵傛偭偰昞旂嵶朎偵庢傝崬傑傟傞丅偙偺IRT1偼嵶朎枌偵嬊嵼偟偰偍傝Fe, Zn, Mn, Cd側偳偺嬥懏僀僆儞傕桝憲偡傞(Eide et al., 1996; Vert et al., 2002; Colangelo and Guerinot, 2004)丅
揝偼媧廂偝傟偨偺偪僯僐僠傾僫儈儞傗僋僄儞巁側偳偺僉儗乕僩偲寢崌偟偰捠摫慻怐偵擖傞丅
偙偺楢摦偟偨揝寚朢墳摎偼basic helix-loop-helix (bHLH) 揮幨場巕偱偁傞FER-LIKE FE DEFICIENCYINDUCED TRANSCRIPTION FACTOR (FIT; Colangelo and Guerinot, 2004)偵傛偭偰惂屼偝傟偰偄傞丅
FIT偼懠偺2偮偺bHLH僞儞僷僋(Yuan et al., 2008) ETHYLENE
INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1
(Lingam et al., 2011)偲嫤憈偟偰偍傝NO偺懚嵼壓偱26僾儘僥傾僝乕儉埶懚惈暘夝偲偄偆東栿屻偺惂屼傪庴偗傞丅偙偺FIT偵埶懚偟側偄揝偺儂儊僆僗僞僔僗偺宱楬偑偁傝偦傟偵偼bHLH100偲bHLH101 (Sivitz et al., 2012)偑娭梌偟偰偄傞偙偲偑摨掕偝傟偨丅
嬤擭Strategy-II怉暔偺墳摎偵帡偨揝寢崌惈僉儗乕僩壔崌暔偲偟偰僼僃僲乕儖壔崌暔傗儕儃僼儔價儞壔崌暔偑傾儔價僪僾僔僗傗Medicago truncatula偐傜暘斿偝傟偰偄傞偙偲偑柧傜偐偵偝傟偰偒偨丅僼僃僯儖僾儘僷僲僀僪傗僼儔價儞崌惉偺戙幱宱楬偼揝妉摼婡峔峔惉偺僐傾堚揱巕偱偁傞FIT, FRO2, IRT1, and AHA2, 偵嫮偔儕儞僋偟偰偄傞丅側偺偱偙傟傜偼傾儔價僪僾僔僗傗M. truncatula偺Strategy-I偺峔惉暘偲傕峫偊傜傟傞丅
変乆偼bHLH僞儞僷僋偱偁傞POPEYE (PYE)偑傾儔價僪僾僔僗偺揝寚朢墳摎斀墳傪億僕僥僀僽偵惂屼偟偰儂儊僆僗僞僔僗傪堐帩偟偰偄傞廳梫側栶妱傪壥偨偟偰偄傞偙偲傪曬崘偟偨丅PYE偼偄偔偮偐偺嬤墢儂儌儘僌偱偁傞IAA-LEUCINE RESISTANT3 (ILR3), bHLH104, bHLH115, 側偳偲憡屳嶌梡偡傞丅埲屻偙傟傜傪PYEL僞儞僷僋偲徧偡傞丅
偙傟傜偺PYELs偼揝寚朢偱up儗僊儏儗僀僩偝傟偰偍傝(Long et al., 2010)丄偦偺偆偪偺堦偮偱偁傞ILR3偼傛偔抦傜偊偰偄傞傛偆側僆乕僉僔儞偲嫟栶偟偨壛悈暘夝傪揝埶懚揑側傗傝曽偱惂屼偟偰偄傞丅偍偦傜偔偼揝僩儔儞僗億乕僞乕椶偑堚揱巕敪尰傪揮幨惂屼偡傞傗傝偐偨偱(Rampey et al., 2006)丅
PYE偲堎側傝丄偁偲偺3偮偺PYEL僞儞僷僋偼戞2偺僞儞僷僋偱偁傞BRUTUS (BTS)偲憡屳嶌梡偟偰偄傞丅BTS偼掅擹搙偺揝偵墳摎偟偰桿摫偝傟偰偍傝丄BTS偺桿摫傪寚幐偝偣傞偲揝寚朢懴惈偑憹壛偟丄wild-type偺怉暔偵斾傋偰崻偺怢挿傗丄崻寳巁惈壔偑憹偡(Long et al., 2010)丅
BTS偼儐僯乕僋側偄偔偮偐偺曐懚僪儊僀儞傪帩偭偰偄傞丅俶枛抂懁偵3屄偺hemerythrin (HHE) 僇僠僆儞寢崌僪儊僀儞傪帩偪丄俠枛抂懁偵偼CHY zinc-finger 僪儊僀儞偲Really Interesting New Gene (RING)僪儊僀儞傪帩偭偰偄傞丅
曐懚偝傟偨HHE僪儊僀儞偵偼left-twisted 4-a-helical bundles偑偁傝丄偙偙偼慳悈惈億働僢僩傪宍惉偟偰偍傝丄偦偙偱O2偑2屄偺揝偲攝埵寢崌偟偰偄傞丅
揝偼HHE僪儊僀儞偺拞偱Glu偲Asp偺僇儖儃僉僔儖婎偲5偮偺His巆婎偲揟宆揑側寢崌傪偟偰偄傞丅偟偐偟懠偺攝埵寢崌偺巇曽傕採埬偝傟偰偄傞(Holmes et al., 1991; Holmes and Stenkamp, 1991; Shu et al., 2012)丅丂BTS僞儞僷僋偺C枛抂嬤朤偺RING僪儊僀儞偺懚嵼偼丄偦偙偑E3 ligase偺擻椡傪桳偡傞偐傕偟傟側偄偙偲傪帵嵈偟偰偄傞丅
僀僱偺BTS偲BTS僆儖僜儘僌偼Oryza sativa Hemerythrin motif-containing RING and Zinc-Finger Protein1 (OsHRZ1)偲OsHRZ2偱偁傞偑揝傪HHE僪儊僀儞偱寢崌偟丄儐價僉僠儞壔擻傪桳偟偰偄傞(Kobayashi et al., 2013)丅偟偐偟BTS偑憡屳嶌梡偡傞揝墳摎惈偺PYEL僞儞僷僋偑E3儕僈乕僛妶惈偺僞乕僎僢僩偱偁傞偐偳偆偐偼挷傋傜傟偰偄側偄丅
偦偙偱変乆偼BTS偼PYEL揮幨場巕傪昗揑偵偟偰RING僪儊僀儞(E3丂ligase)傪捠偠偰偺26S proteasome-宱桼偺暘夝偵娭梌偡傞揝儂儊僆僗僞僔僗偺僱僈僥僀僽儗僊儏儗乕僞乕乮晧偺挷愡場巕乯偱偁傠偆偲偄偆壖愢傪棫偰偨丅
偙偺E3儕僈乕僛妶惈偼BTS偺埨掕惈偺曄壔偵埶懚偟偰偍傝丄偦偺埨掕惈偼HHE僪儊僀儞偵塭嬁偝傟丄偟偨偑偭偰揝
偺棙梡惈偺梙傜偓乮曄摦乯偵墳摎偟偰偄傞丅
変乆偼BTS偺敪尰傪怉暔偺敪払抜奒傪捠偟偰條乆側慻怐偱扵嶕偟丄揝寚朢墳摎偲惗怋惉挿偵偍偗傞BTS敪尰偺曄摦偺堄枴偵偮偄偰傕暘愅偟偨丅
摨帪偵変乆偼BTS偺HHE僪儊僀儞傪夘偟偨揝寢崌擻傪挷傋偨丅揝偑擹搙埶懚揑偵BTS偺埨掕惈偵塭嬁偟丄HHE僪儊僀儞偺摿堎揑傾儈僲巁巆婎偑偙偺抈敀偺埨掕惈偵寛掕揑偱偁傞偙偲傪徹柧偟偨丅
変乆偼偝傜偵BTS僞儞僷僋偲PYEL僞儞僷僋偺憡屳嶌梡偑RING僪儊僀儞偵埶懚偡傞帠丄傑偨BTS偑彮側偔偲傕2偮偺PYEL僞儞僷僋偑26S proteasome傪宱桼偡傞暘夝傪彆偗傞偙偲傪尒弌偟偨丅
寢榑偲偟偰変乆偺幚尡寢壥偵傛偭偰丄BTS偑丄摿偵bHLH揮幨場巕傪惂屼偡傞偙偲偵傛偭偰丄怉暔偵偍偗傞揝儂儊僆僗僞僔僗偺堐帩偵廳梫側栶妱傪桳偡傞偙偲偑柧傜偐偵側偭偨丅
丂丂丂
埲壓恾偺愢柧
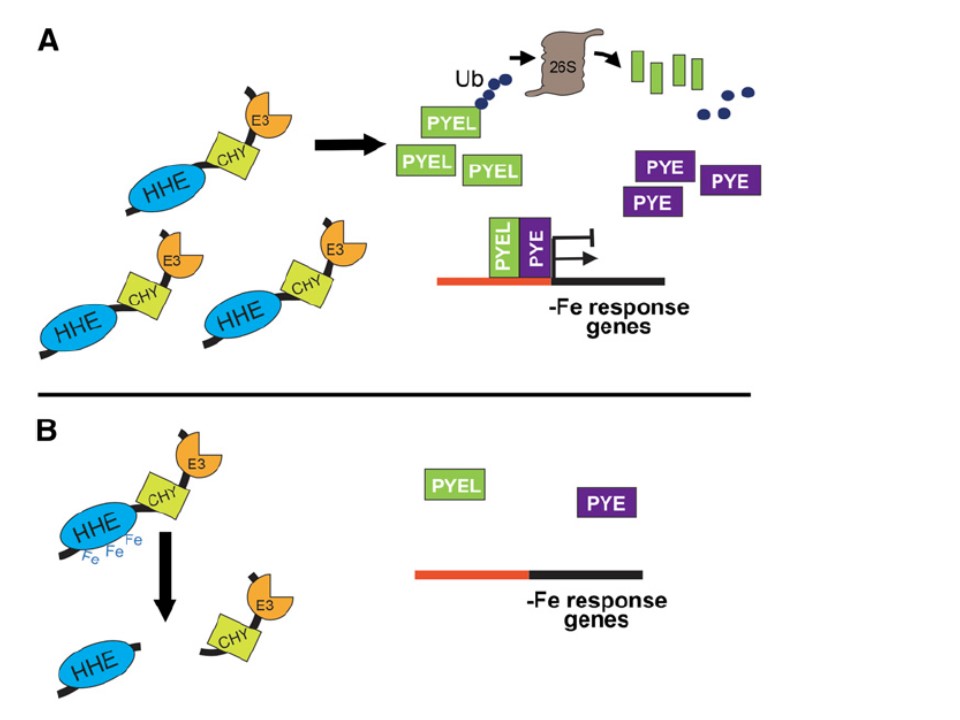
BTS僞儞僷僋偺埨掕惈偲偦偺嶌梡儌僨儖
A. BTS僞儞僷僋偼丄bHLH宆偺揮幨場巕偱偁傞PYE偲PYEL偲楢摦偟偰揝寚朢偱揮幨桿摫偝傟傞丅掅擹搙偺揝塰梴忦審壓偱偼BTS僞儞僷僋偼傛傝埨掕偱偁傝PYEL/PYE偺惂屼妶惈傪E3儔僀僎乕僗妶惈偱惂屼偟偰偄傞丅偮傑傝PYEL/PYE宱桼偺怉暔偺揝寚朢墳摎傪挷惍偟偨傝旝挷惍偟偨傝偟偰偄傞丅
B. 揝偑嫙梌偝傟傞偲丄BTS丄PYE丄PYEL偺揮幨儗儀儖偱偺桿摫偑掅壓偡傞丅揝擹搙偺忋徃偑HHE僪儊僀儞偱専抦偝傟傞偲BTS僞儞僷僋偺宍忬偲埨掕惈偑曄壔偡傞丅偦偺墳摎偺寢壥BTS僞儞僷僋偺暘夝偑婲偙傝丄偦偺屻偺嵶朎偺揝墳摎惈E3儔僀僎乕僗妶惈偑掅壓偡傞丅
BTS偺嶌梡儌僨儖