乽IRON丂MAN乿 偼怉暔偺揝桝憲傪僐儞僩儘乕儖偡傞曃嵼(儐價僉僞僗側乯儁僾僠僪僼傽儈儕乕偱偁傞
IRON丂MAN 偼怉暔偺揝桝憲傪僐儞僩儘乕儖偡傞儐價僉僞僗側儁僾僠僪僼傽儈儕乕偱偁傞
IRON MAN, a ubiquitous family of peptides that丂control iron transport in plants
Louis Grillet, Ping Lan, Wenfeng Li, Girish Mokkapati & Wolfgang Schmidt
doi: https://doi.org/10.1101/351635
丂
偙偺榑暥偼3擭3偐寧偠傜偝傟偨偺偪偵 Nture plants偵宖嵹偝傟偨丅
傑偊偑偒乮屻敿晹暘偺彺栿乯
抧忋晹偐傜偺僔僌僫儖偑崻偺揝媧廂傪惂屼偟偰偄傞丄偲偄偆偙偲偼夁嫀20擭埲忋
傕偺娫丄揝傪廤愊偡傞宍幙傪桳偡傞戝摛偺乭倓gl乭曄堎姅傪梡偄偨愙偓栘朄偱幚徹偝傟偰偄偨丅
傾儔價僪僾僔僗偱偼丄frd3傗opt3偺曄堎姅偱偼丄偦傟偧傟揝偺崻偐傜梩傊偺堏峴偺寚幐偲丄揝偺巘娗傊偺愊傒崬傒偺寚幐偐傜偔傞丄揝偺媧廂懀恑偲偄偆椺偐傜丄偦偺傛偆側僔僌僫儖偑旛傢偭偰偄傞徹嫆偑偁偭偨丅
偟偐偟丄乽梩偺揝偺忬懺偺忣曬傪崻偵揱偊傞挿嫍棧桝憲僔僌僫儖偺惈幙乿偮偄偰偼丄塱偄偁偄偩怉暔偺揝塰梴尋媶偱偼柪媨擖傝偱偁偭偨丅
拏慺寚朢偺崻偱偼C枛抂偵15傾儈僲巁巆婎傪僐乕僪偟偨儁僾僠僪(CEPs)偑leucinerich repeat receptor kinase CEPR傪妶惈壔偡傞偙偲偑抦傜傟偰偄傞丅
CDPR偼偍偦傜偔巘娗偵嬊嵼偡傞 僋儔僗III 僞僀僾偺glutaredoxin CEPD1
傪儕儞巁壔偟丄偦傟偑偦偺屻丄巘娗偐傜撪旂偵堏峴偡傞擻椡傪恎偵拝偗傞丅
偳偺傛偆偵偟偰CEPD1偑 徤巁媧廂僩儔儞僗億乕僞乕 乬NRT2.1乭偺敪尰偺堷偒嬥傪堷偔偺偐偼傑偩暘偐偭偰偄側偄丅
偙偙偵変乆偼巘娗偱敪尰偟偰偄傞丄偍偦傜偔巘娗偺拞傪摦偄偰偄傞僔僌僫儖偱傾儔價僪僾僔僗偺揝媧廂傪僐儞僩儘乕儖偟偰偄傞怴偟偄儁僾僠僪僼傽儈儕乕傪敪尒偟偨偙偲傪偺傋傞丅偙傟偼偍偦傜偔旐巕怉暔Magnoliophyta娫偵嫟捠偺揝僔僌僫儖惉暘偱偁傞丅
偙偺僼傽儈儕乕偼17傾儈僲巁巆婎偺僐儞僙儞僒僗儌僠乕僼傪C枛抂偵娷傓傕偺偱偁傝丄偙偺儌僠乕僼偼旐巕怉暔娫偱崅搙偵曐懚偝傟偰偍傝丄搚忞偐傜偺揝媧廂偺堷偒嬥偲側傞偺偵昁梫偐偮廫暘側傕偺偱偁傞丅
丂丂丂丂丂
(梫栺乯
揝偼怉暔廩暘偵嫙媼偝傟側偄応崌偼怉暔偺惉挿傗丄廂検傗丄塰梴揑昳幙偵怺崗側塭嬁傪梌偊傞丅
抧忋晹偺僔僌僫儖偵傛偭偰崻偺揝媧廂偑惂屼偝傟偰偄傞偲偄偆偙偲偑愙偓栘朄偱揝廤愊宯摑偺戝摛曄堎姅dgl偵傛偭偰帵偝傟偰偐傜20擭埲忋宱夁偟偨丅儔價僪僾僔僗偱偼偦偺傛偆側僔僌僫儖偺懚嵼偑丄崻偐傜抧忋晹傊偺揝偺堏峴擻椡偺寚幐傗丄巘娗傊偺揝偺愊傒壓傠偟擻椡偑寚幐偡傞偲偄偆偙偲偼丄偦傟偧傟曄堎姅frd3偲 曄堎姅opt3偱帵偝傟偰偄偨丅偟偐偟梩偺揝偺懚嵼忬懺傪崻偵揱偊傞挿嫍棧桝憲忣曬偺幚懱偑壗偱偁傞偐偼揝塰梴尋媶偺挿偄娫偺撲偱偁偭偨丅
丂丂丂丂
丂丂椺偊偽丄拏慺尮偑愨偨傟偨崻偱偼C乗枛抂偺15傾儈僲巁巆婎偺僼傽儈儕乕(CEPs)偑leucine-rich repeat receptor kinase CEPR傪妶惈壔偡傞丅CEPR偼巘娗偵嬊嵼偡傞僋儔僗III glutaredoxin CEPD1傪儕儞巁壔偟丄偙傟偑寢壥揑偵巘娗偐傜弌偰撪旂偵堏峴偡傞丅偳偺傛偆偵偟偰CEPD1偑NRT1.1, NRT2.1 and NRT3.1側偳偺徤巁媧廂僩儔儞僗億乕僞乕偺敪尰偺堷偒嬥傪堷偔偺偐偼丄傑偩撲偱偁傞偑丅
丂丂
丂丂偙偺榑暥偱偼丄巘娗偱敪尰偟偰偄偰懡暘傾儔價僪僾僔僗偺揝媧廂偺巘娗傪堏摦偡傞僔僌僫儖偱偁傝丄偙傟偑偨傇傫旐巕怉暔偺嫟捠惉暘偱偁傞偲巚傢傟傞怴偟偄儁僾僠僪僼傽儈儕乕傪敪尒偟偨偺偱曬崘偡傞丅
変乆偼偙偺曬暥偱旐巕怉暔偺側偐偱悢懡偔偺崅搙偵懡條側儁僾僠僪椶偺C-枛抂傾儈僲巁巆婎偵嫟捠偺儌僠乕僼(IRON MAN; IMA)偑曐懚偝傟偰偄傞偑丄偙傟偑怉暔偺揝妉摼偵偲偭偰昁恵偱偁傞偙偲傪帵偡丅
丂丂
丂丂壓婰偺IMA攝楍傪傾儔價僪僾僔僗偱夁忚敪尰偝偣傞偲崻偱偺揝媧廂堚揱巕孮傪桿摫偟庬巕傪偼偠傔偲偡傞怉暔偺慡慻怐偱揝偲儅儞僈儞偺拁愊偑婲傞丅傾儔價僪僾僔僗僎僲儉偺IMA堚揱巕敪尰傪梷惂偡傞偲揝媧廂偑柍岠偵側傝怺崗側僋儘儘僔僗偑婲偙偭偨丅偙偺帪揝嫙梌傪憹壛偝偣傞偐IMA1堚揱巕傪夁忚敪尰偝偣傞偲栰惗宆偺昞尰宆偵暅婣偟偨丅
丂丂
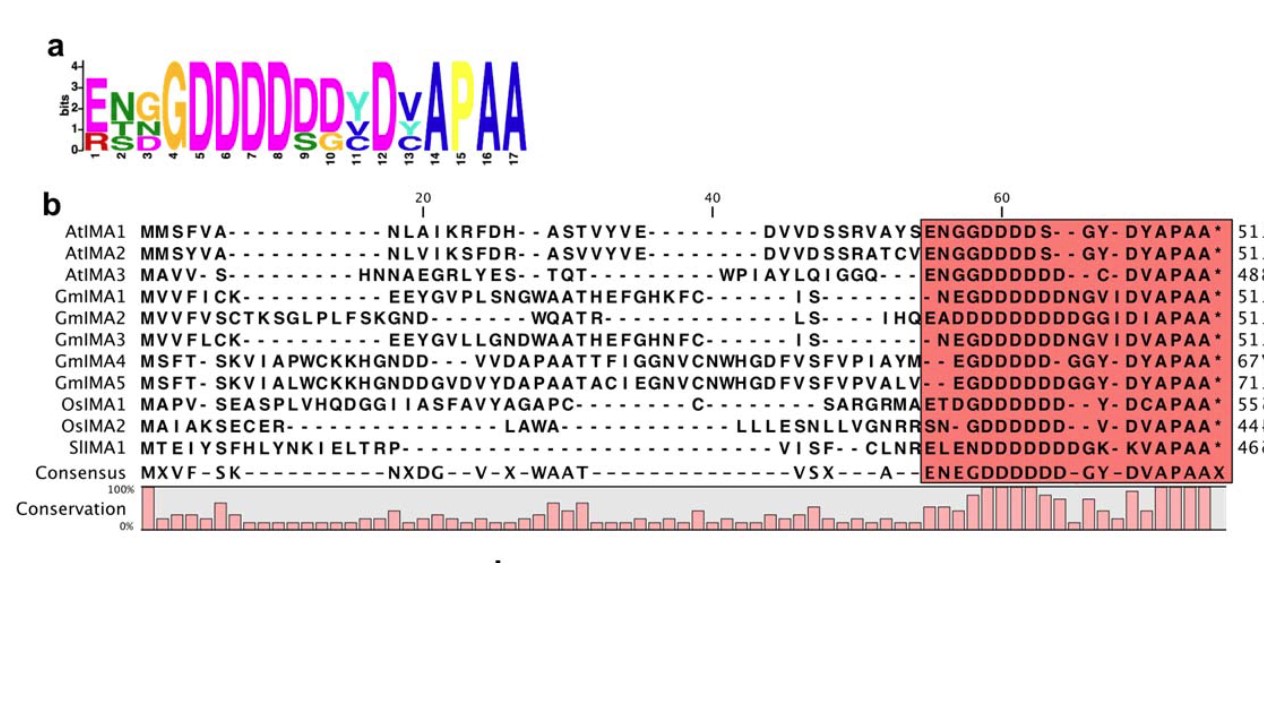
丂丂At1g47400偼 G-D-D-D-D-x(1,3)-D-x- A-P-A-A偲偄偆嫟捠偺傾儈僲巁攝楍傪帩偭偰偍傝丄偙偺攝楍傪丂IRON MAN (IMA)丂偲柤晅偗偨. 傾儔價僪僾僔僗偺僎僲儉偵偼8偮偺 IMA堚揱巕偑偁傝丄偦傟傜偼偡傋偰揝偺嫙梌偵墳摎偟偨丅 摿偵AtIMA1 (At1g47400)丄 AtIMA2 (At1g47395) 丄 AtIMA3 (At2g30766)偺3偮偼梩偲崻偱揝寚朢偱嫮偔敪尰偟偨丅
丂丂丂
丂丂偙偺曐懚攝楍椞堟偑IMA僞儞僷僋偺嶌梡晹埵偱偁傞偲偄偆偙偲傪妋偐傔傞偨傔偵丄IMA1偺17C-枛抂傾儈僲巁巆婎偺忋棳偵奐巒僐僪儞傪偮偗偰夁忚敪尰偝偣偨丅捠忢偺忦審壓偱偙偺僐儞僗僩儔僋僩偼IMA1偺夁忚敪尰宯摑(Ox line)偲摨條側嶰壙揝娨尦峺慺妶惈傪帵偟偨丅摨條偵偟偰嶌惉偟偨傎偐偺7偮偺崌惉IMA傕嫮庛偼偁傟摨條偺孹岦傪帵偟偨偺偱丄IMA攝楍偼堚揱巕娫偱偺儕僟儞僟儞僔乕乮忕挿惈乯傪帩偮偲巚傢傟偨丅
丂丂丂
丂丂偙傟傜偺寢壥偐傜IMA1偺IMA儌僠乕僼偼崻偱偺揝媧廂偵昁梫偐偮廫暘偱偁傞偙偲偑暘偐偭偨丅IMA1偼庡偲偟偰巘娗偱丄偳偪傜偐偲偄偊偽梩偱丄惓媡愙偓栘朄偵傛傞偲抧忋晹偺MA1儁僾僠僪椶偼崻偺揝媧廂傪億僕僥僀僽乮惓乯偵惂屼偟偰偄偨丅IMA偺儂儌儘僌偼揝偺懚嵼忬懺偵墳摎偟丄堎庬娫偱傕敪尰偑嶌摦偟偨丅
丂丂丂丂丂
丂丂IMA偼棨惗怉暔偺揝妉摼偲揝偺嵶朎撪儂儊僆僗僞僔僗偵杮幙揑側儁僾僠僪僼傽儈儕乗偲峫偊傜傟傞丅
丂丂
乮幚尡曽朄側偳棯乯丂丂
丂丂丂
丂丂丂丂丂丂
乮榑媍乯
杮榑暥偱丄変乆偼崅搙偵揝墳摎惈偺怴婯側儁僾僠僪僼傽儈儕乕傪摨掕偟偨偑丄偙偺C-枛抂傾儈僲巁攝楍偺儌僠乕僼偼揝媧廂偵偲偭偰寚偔偙偲偺偱偒側偄傕偺偱偁傞丅
偙偺IMA儁僾僠僪椶傪僐乕僪偡傞堚揱巕傪僨乕僞儀乕僗偐傜専嶕偡傞偲丄棙梡壜擻側僨乕僞偡傋偰偺旐巕怉暔偵懚嵼偡傞偺偱丄壴偑嶇偔怉暔偵儌僠乕僼偑曐懚偝傟偰偄傞偲峫偊傜傟傞丅
丂丂
揝寚朢壓偱嫮偔敪尰偟偰偄傞偵傕偐偐傢傜偢摉弶偼IMA儁僾僠僪偑堦偮偺僼傽儈儕乕偱偁傞偙偲偑擣幆偱偒側偐偭偨丅偦傟偼丄偄偔偮偐偺惂栺偺偨傔偵嫟桳偡傞僐儞僙儞僒僗攝楍偲偟偰摨掕偡傞偙偲偑偱偒側偐偭偨偨傔偱偁傞丅
丂丂丂
IMA儁僾僠僪傪BRAST専嶕偵偐偗傞偲N枛抂攝楍偑崅昿搙偵曄摦偟偰偄傞偙偲丄1偮偺Asp巆婎偑怢傃偰偄傞掱搙偺暋嶨惈偑専嶕傾儖僑儕僘儉偺儌僠乕僼傪暍偄塀偡偙偲丄儌僠乕僼偺拞娫偵懚嵼偡傞崅搙偺曄摦偡傞傾儈僲巁椶偵傛傞嬻寗丄ORF僒僀僘偑俀俁乣俉俇傾儈僲巁偵傑偱戝偒偔曄摦偟偰偄傞偙偲側偳丄條乆側専嶕偵娭傢傞朩奞梫場偑偁偭偨丅
丂丂丂
柺敀偄偙偲偵丄晹暘揑側IMA儌僠乕僼偼丂alphaproteobacteria 椺偊偽AgrobacteriumTumefaciens偺傛偆側怉暔偲憡屳嶌梡偟偰揝妉摼偡傞庡梫側儗僊儏儗乕僞乕乮惂屼場巕乯偱偁傞rirA偺C-枛抂偵傕丄Streptomyces 僶僋僥儕傾偺TonB乮揝僔僨儘僼僅傾儕僙僾僞乕乯偵傕尒弌偝傟偨丅
丂丂
傾儔價僪僾僔僗偺僎僲儉偵偼俉偮偺IMA堚揱巕偑娷傑傟偰偍傝丄偦偺偆偪偺6偮偑non-coding RNAs (IMA3–8)偲巚傢傟傞丅偙傟傜偼丄
彫偝側僒僀僘偺ORF偲orthologue偑懚嵼偟側偄偲偄偆BLAST夝愅偐傜梊應偟偨傕偺偱偁傞丅
丂丂丂
IMA偺僐儞僙儞僒僗傾儈僲巁攝楍偼崅搙偵曐懚偝傟偰偄傞偙偲丄儁僾僠僪N枛抂偺曐懚偝傟偰偄側偄攝楍傪庢傝彍偄偨応崌偵傕IMA儌僠乕僼偑嶌摦偡傞帠側偳偼丄IMA倱偑儁僾僠僪儗儀儖偱嶌梡偡傞偙偲傪嫮偔巟帩偟偰偄傞丅
偙偺壖愢偼IMA1偲IMA3儊僢僙儞僕儍乕RNA傪genome-wide ribosome profiling surveys偱東栿偟偰娤嶡偝傟偨偙偲偵傛偭偰傕朤徹偝傟偨丅
丂丂丂
偟偐偟丄怉暔偵撪嵼偡傞IMAs偼峈懱偱傕幙検暘愅偱傕専弌偝傟側偐偭偨丅
丂丂
偙偺偙偲偼儁僾僠僪偑杮棃晄埨掕側偺偐丄揮幨嶻暔偐傜偺東栿儗儀儖偑掅偄偨傔偐丄偦偺椉幰側偺偐傕偟傟側偄丗傾僗僷儔僊儞巁巆婎偑怢挿偟偰偄傞偙偲偑丄東栿偵晧偺僀儞僷僋僩傪梌偊偰偄偨丅
丂丂丂
IMA倱偑儁僾僠僪偲偟偰摥偔偺偩偲偄偆偙偲偼丄IMA1:EYFP梈崌僞儞僷僋偑怉暔偺拞偱FCR(嶰壙揝娨尦峺慺妶惈)偺憹壛偝偣傞偙偲傪娤應偟偨偲偄偆捈愙揑側徹柧偵傛傞傕偺偱偁傞丅
丂丂丂
IMA倱偑揝妉摼堚揱巕傪惂屼偟偰偄傞偲偄偆丄徻嵶側暘巕儊僇僯僘儉偼傑偩柧傜偐偱偼側偄偑丄Fe2+偑寢崌偡傞偙偲偵傛偭偰IMA儁僾僠僪偑埨掕壔偟丄偦傟偑揝媧廂傪惂屼偟偰偄傞偺偩偲巚傢傟傞丅
丂丂丂
傾僗僷儔僊儞巁偺傛偆側晧偵僠儍乕僕偟偨巆婎偼悢懡偔偺僞儞僷僋偲攝埵寢崌偟偰偄傞丅IMA儌僠乕僼偺Asp偺怢傃暘偼嬥懏偲寢崌偡傞惈幙偑偁傞偩傠偆偲峫偊偰幚尡偟偨偑丄梊憐捠傝IMA儁僾僞僀僪偑Fe2+傗懠偺嬥懏偲寢崌偡傞偙偲傪帵偟偨丅
丂丂
偙傟傜偺寢崌僒僀僩偑朞榓偝傟傞偲儁僾僠僪偑捑揳偡傞堷偒嬥偵側偭偨丅
丂丂丂
IMAs偼庡偵掅揝嫙媼忦審壓偱敪尰偡傞偺偱丄乽IMAs偑晄埨掕偱偁傞偲偄偆偙偲偑乮崻偺乯揝媧廂婡峔偵僱僈僥僀僽僼傿乕僪僶僢僋偵嶌梡偡傞丄偮傑傝丄揝廫暘忦審壓偱偺巘娗偺揝偵傛偭偰偙偺僼傿乕僪僶僢僋儊僇僯僘儉偼堷偒嬥傪堷偐傟傞偺偱偁傞乿偲偄偆壖愢傪棫偰偨丅
丂丂丂
偙偺偙偲偼夁忚敪尰宯偱傕丄偛偔傢偢偐偟偐EYFP:IMA1梈崌僞儞僷僋偑廤愊偟側偄偙偲丄撪惗IMA儁僾僞僀僪偺専弌偑崲擄側偙偲丄栰惗宆偺愙偓曚偐傜偺IMAs偺岠壥偑ima8x 戜嵗偺FCR妶惈偵懳偟偰斾妑揑揔搙側掱搙偱偁傞偙偲側偳傪傛偔愢柧偱偒偰偄傞丅
丂丂
偄偔偮偐偺旐巕怉暔偵偍偄偰揝寚朢偵墳摎偡傞IMA傪僐乕僪偡傞堚揱巕孮偼敪尰偑幚幙揑偵桿摫偝傟傞偙偲偐傜丄IMA僼傽儈儕乕偺儁僾僠僪偺嶌梡偑揝儂儊僆僗僞僔僗偺揰偐傜傕庬娫偱曐懚偝傟偰偄傞偙偲傪帵嵈偟偰偄傞丅
丂丂
IMAs堚揱巕敪尰偼摵乮Cu乯傑偨偼垷墧(Zn)寚朢偱偼惂屼偝傟偰偍傜偢丄偄偔偮偐偺椙偔妋棫偝傟偨揝乮Fe乯偵摿堎揑側惂屼堚揱巕敪尰偲崅偄憡娭偑偁傞丅
丂
IMA儌僠乕僼傪夁忚敪尰偝偣傞偲怉暔偑揔摉側揝塰梴忦審壓偱嶌摦偟偰偄傞揝媧廂梷惂傪柍岠偵偟偰丄崻偺嵶朎偺揝寚朢墳摎斀墳偺堷偒嬥傪堷偒丄崅恊榓惈揝僩儔儞僗億乕僞乕偱偁傞IRT1偺戞堦傗戞擇偺婎幙偱偁傞丂Fe2+, Mn2+, Zn2+ 側偳偺崻偵偍偗傞娷検傪崅傔丄側偍偐偮抧忋晹偱偺娷検傕崅傔傞丅
丂丂
懠曽偱偼丄ima8x曄堎姅偺偡傋偰偺IMA堚揱巕傪僒僀儗儞僔儞僌偡傞偲揝嫙媼偺媫寖側寚懝偲丄揝寚朢墳摎擻偑幐傢傟偰丄屚巰偵帄傞丅
丂丂
偱偁傞偐傜丄偄傑傗IMAs偼嵶朎偺揝儂儊僆僗僞僔僗偺摑崌惉暘傪戙昞偡傞傕偺偱偁傞偲峫偊傜傟傞丅偙傟偼傾儔價僪僾僔僗偺傛偆側娨尦宆愴棯乮Strategy-I僞僀僾乯傪慖戰偟偨庬偵尷掕偝傟傞傕偺偱偼側偄偩傠偆丅
変乆偺僨乕僞偼偝傜偵IMA儁僾僠僪偺儗儀儖乮偙偙偱偼奒憌偲栿偡傋偒偐乯偑庬摿堎揑揝妉摼婡峔偺忋棳偵嶌梡偟偰揝妉摼偺巜椷搩偵埵抲偡傞偙偲傪帵偟偰偄傞丅
丂
ima8x曄堎姅偺嫮偄昞尰宆傪梡偄偨幚尡偱偼丄婡擻惈IMAs偑丄揝廫暘忦審壓偱傕揝寚朢忦審壓偱傕丄嵶朎偺揝儂儊僆僗僞僔僗偵偲偭偰昁恵偱偁傞偙偲傪帵偟偰偄傞丅
IMA偺夁忚敪尰宯楍(IMA Ox lines)偺夁忚揝娷検昞尰宯偼丄抧忋晹偐傜崻傊偺僔僌僫儖傪寚懝偟偨揝夁忚廤愊曄堎姅偱偁傞opt3傗戝摛偺曄堎姅 dgl傪楢憐偝偣傞丅偙偺偙偲偼IMAs偑慻怐娫惂屼偵娭傢傞揝媧廂偺懀恑僔僌僫儖偺栶妱傪扴偆偲偄偆悇掕傪巟帩偟偰偄傞丅
丂丂丂
IMA偺婡擻偼丄慡恎揑拏慺僔僌僫儖偲偟偰抦傜傟傞CEPD1/CEPD2偺栶妱偲椶帡偟偰偄傞丅偙偺偙偲乮傪晛曊壔偡傞偲丠乯偼丄怉暔偱偼儁僾僠僪椶偑搚忞偐傜柍婡塰梴慺傪妉摼偡傞偲偒偺旝挷惍傪梫媮偡傞偲偒偺曇惉嬈柋偲偄偆寛掕揑側栶妱傪扴偭偰偄傞偺偐傕偟傟側偄丅
偙偺榑暥偲傎傏摨帪婜偵壀嶳戝妛偐傜埲壓偺忣曬偑敪怣偝傟偨丅偙偺榑暥偲摨偠儁僾僠僪偺敪尒偱偁傞丅
尋媶惉壥 > 怉暔偺揝墳摎偵娭傢傞怴婯抁嵔儁僾僠僪場巕傪摨掕
怉暔偺揝墳摎偵娭傢傞怴婯抁嵔儁僾僠僪場巕傪摨掕
[挊幰]丂Hirayama, T., Lei, G.J., Yamaji, N., Nakagawa, N., Ma, J.F.
[榑暥僞僀僩儖]丂The putative gene FEP1 regulates iron deficiency response in Arabidopsis
[宖嵹榑暥]丂Plant Cell Physiology (2018) 59: 1739-1752
https://academic.oup.com/pcp/article/59/9/1739/5056076
[撪梕徯夘]丂揝偼惗暔偵昁恵偺尦慺偱偡丅搚忞拞偵娷傑傟傞揝偺懡偔偼棙梡崲擄側晄梟惈偺嶰壙揝偱偡偑丄怉暔偼搚忞偐傜揝傪岠棪傛偔媧廂偡傞偙偲偑偱偒傑偡丅揝偼昁恵偱偡偑偦偺夁忚偼桳撆偱丄揝媧廂丒桝憲偼尩枾偵惂屼偝傟偰偄傑偡丅偙傟傑偱偺尋媶偐傜丄僔儘僀僰僫僘僫偱偼丄揝寚朢忬嫷偱揮幨場巕FIT, bHLH38, 39側偳偑丄揝媧廂丒桝憲偵娭傢傞堚揱巕孮傪妶惈壔偟丄堦曽BTS偲柤晅偗傜傟偨場巕偑丄揝媧廂桝憲堚揱巕孮偺敪尰傪梷惂偟丄揝偺峆忢惈堐帩偵婑梌偡傞偙偲偑帵偝傟偰偄傑偡丅偟偐偟丄怉暔偑偳偺傛偆偵揝寚朢傪擣幆偡傞偺偐丄傑偨偦偺忣曬偑偳偺傛偆偵bHLH39傗FIT傪妶惈壔偡傞偺偐徻嵶偼晄柧偱偡丅偝傜偵丄揝傪媧廂偡傞崻偲揝傪徚旓偡傞抧忋晹偺娫偱偺忣曬岎姺傗屄懱慡懱偱揝寚朢墳摎傪摑屼偡傞巇慻傒偑憐掕偝傟偰偄傑偡偑丄偦偺幚懺偼柧傜偐偱偼偁傝傑偣傫丅崱夞丄怴婯偺47傾儈僲巁偐傜側傞抁嵔儁僾僠僪FEP1偑僔儘僀僰僫僘僫偺揝寚朢墳摎偱廳梫側栶妱傪扴偭偰偄傞偙偲傪柧傜偐偵偟傑偟偨丅崱夞偺尋媶偐傜丄FEP1偼bHLH39傗FIT側偳偺揮幨場巕傛傝忋棳偱婡擻偡傞偙偲偑傢偐傝傑偟偨丅傑偨丄fep1曄堎姅偺夝愅側偳偐傜FEP1偑慻怐娫偺墦嫍棧忣曬揱払偵娭傢傞壜擻惈傕帵嵈偝傟傑偟偨丅崱屻丄FEP1傪婲揰偵揝寚朢墳摎婡峔偺夝柧偑恑傓偙偲偑婜懸偝傟傑偡丅偙偺尋媶偼丄慡崙嫟摨棙梡嫟摨尋媶嫆揰帠嬈偵傛傝僒億乕僩偝傟傑偟偨丅乮暥愑丂娐嫬墳摎婡峔尋媶僌儖乕僾丒暯嶳 棽巙乯
揝寚朢偵惓偵墳摎偡傞IMA嫟捠攝楍