ニコチアナミン合成遺伝子(NAS)導入でバイオ強化したパン小麦の複数年圃場試験評価
以下の論文は、受理(雑誌名未公開)されているが試読で公開されているものである。イネのニコチアナミン合成酵素遺伝子(NAS2)を小麦に導入した、オーストラリアの3カ所でのフィールド試験の結果である。読んでいて実に堅実な、多角的で示唆的なdiscussionがなされていることに感銘を受けたので紹介する。
きちんとした遺伝子組み換え圃場でやっていることが写真からわかる。
オーストラリアとアメリカのグループの共同研究である。
この研究で最も顕著な成果はNAS2を遺伝子導入した小麦は栽培環境を問わず穀粒中のニコチアナミンとデオキシムギネ酸含有量が増加しているという顕著な事実である。これが人体でもパンの鉄の消化吸収に効果があるのではないかという、重要な次のテーマを提案している。
追記:その後PLANT Journal誌に
掲載された。
https://onlinelibrary.wiley.com/doi/10.1111/tpj.15623
ニコチアナミン合成遺伝子(NAS)導入でバイオ強化したパン小麦の複数年圃場試験評価
Multi-year field evaluation of nicotianamine biofortified bread wheat
Jesse T. Beasleya , Julien P. Bonneaua , Laura T. Moreno-Moyanoa , Damien L. Callahanb , Kate S. Howellc , Elad Takod , Julian Taylore , Raymond P. Glahnf , Rudi Appelsc , Alexander A. T. Johnsona*
aSchool of BioSciences, The University of Melbourne, Victoria 3010, Australia bSchool of Life and Environmental Sciences, Deakin University, Victoria 3125, Australia cSchool of Agriculture and Food, The University of Melbourne, Victoria 3010, Australia dDepartment of Food Science, Cornell University, Stocking Hall, Ithaca, NY, 14853-7201, USA eSchool of Agriculture, Food and Wine, University of Adelaide, Glen Osmond, South Australia 5064, Australia fRobert W. Holley Center for Agriculture and Health, USDA-ARS, Ithaca, New York 14853, USA
(要約)
パンコムギ(Triticum aestivum L.)の鉄および亜鉛のバイオ強化を目指した従来の育種は、これらの形質に関する遺伝的変異が少なく、穀物の鉄および亜鉛濃度と収量の間に負の相関があることが妨げとなってきた。我々は、イネのニコチアナミン合成酵素 2 (OsNAS2) 遺伝子を構成的に発現させ、2 種類の金属キレーターであるニコチアナミン (NA) と 2'-deoxymugineic acid (DMA) の生合成を促進する遺伝子工学をパンコム ギに適用し、全粒粉および白粉中の鉄および亜鉛の濃度を増加させることに成功した。本報告では、低コピーCE-OsNAS2小麦品種(CE-1)の3年間にわたる多地点限定圃場試験(CFT)評価について述べる。CE-1の全粒粉、白粉および白パン中のNA、DMA、鉄および亜鉛の濃度が高く、a null segregant(NS:非分離群)対照に対してCE-1の白粉の鉄の利用性が高いことを明らかにした。農作物および穀物栄養形質の多環境モデルにより、穀物収量と穀物の鉄、亜鉛および総タンパク質濃度は負の相関を示したが、穀物収量と穀物のNAおよびDMA濃度には相関がなかった。白粉の鉄の生物学的利用能は白粉のNA濃度と正の相関があり、小麦の鉄の生物的強化においてNAキレート鉄がターゲットとされるべきことが示唆された。
(まえがき)
小麦は、パン用小麦(Triticum aestivum L.)とデュラム小麦(Triticum durum L.)の2種類が主で、他のどの作物よりも多くの土地で生産され、パン、麺、パスタなど様々な食品に加工されて人間の1日の摂取カロリーの〜20%を供給している。小麦の生産量は2017年に7億7000万トン(MT)を超え、同年には米(Oryza sativa L.)を抜いてトウモロコシ(Zea mays L.)に次いで2番目に生産量の多い作物となった(FAOSTAT, http://www.fao.org/faostat)。MENAP(中東、北アフリカ、アフガニスタン、パキスタン)地域の国々のように、小麦の一次生産があまり発達していない国では、小麦の消費は1日のカロリー摂取量の40%以上を占め、人間の鉄(Fe)および亜鉛(Zn)不足の高い普及率と一致する。しかし、 小麦の鉄および亜鉛のバイオフォーティフィケーション(生物的強化)は、ゲノム資源の不足、遺伝子型(G)x 環境(E)効果の大きさ、いくつかの環境条件下での穀物収量との負の関連によって妨げられてきた。
過去 20 年間の国際的な育種活動により、Triticum dicoccoides 野生品種の穀物微量栄養素の変異を利用し、合成六倍体の開発によって、穀物 Zn が最大 1.3 倍に増加したエリート耐病性と高収量性の生物的強化小麦が開発された。従来、小麦の鉄分強化のための育種はあまり成功しなかったが、NAM-B1(通称Gpc-B1)の機能的対立遺伝子(この遺伝子は止め葉の成熟と老化とFe とZnの葉から種子への転流を制御する)を選抜することによって、最大1.2倍の穀粒鉄分の増加を達成した。
従来の育種による穀物の生物的強化は、フィチン酸(イノシトール六リン酸)、ポリフェノール、繊維などの、人間の腸内での鉄と亜鉛の吸収(バイオアベイラビリティ)を阻害する食品化合物の存在によっても制限される。
小麦粒に含まれる鉄および亜鉛の大部分(70-80%)は、胚乳の糠層などの粒外組織にあるフィチン酸含有グロボイドと共局在しており、したがって全粒粉中の鉄および亜鉛濃度が増加しても、小麦の消費者の食事性微量栄養素吸収を改善しない可能性がある。小麦の内胚乳に含まれる限られた鉄および亜鉛(20-30%)はフィチン酸と共局在化しないため、より生物学的に利用できる可能性がある。現代の植物バイオテクノロジーにより、研究者は胚乳への負荷に対する生理学的障害を克服し、白小麦粉中の鉄および亜鉛の濃度を増加させることができるようになるはずである。
作物の栄養品質や加工形質を改善するために遺伝子組換え(GM)を利用することは、最近、第二世代 GM と呼ばれ、後進国の人々の健康に役立つ大きな可能性を秘めている。穀物への鉄と亜鉛の取り込みは、金属特異性の高いトランスポーターとキレート剤によって制御されている。小麦の場合、鉄と亜鉛はすべて茎葉を経て、維管束と核突起からなる穀粒領域である腹溝に入り、その後、鉄はアリューロンに、亜鉛は胚と縦溝の組織に再分配される。
アリューロンでは、鉄は液胞鉄輸送体(VIT)遺伝子によって細胞液胞に貯蔵される。また、小麦TaVIT2遺伝子を胚乳特異的高分子量グルテニンプロモーター(HMW)の制御下で発現すると、アリューロンから胚乳に鉄が再配分され、全粒の鉄含量を増加させずに白粉の鉄濃度を2倍にできる。
師管の鉄は、高等植物において鉄、亜鉛、その他の遷移金属をキレートする非タンパク質アミノ酸であるニコチアナミン(NA)とほとんど結合しており、穀類において内因性鉄キレート剤である2'-デオキシムゲン酸(DMA)の生合成前駆体となり、根圏に分泌されてフィトシデロフォアとして鉄をキレートする。白小麦粉中の鉄の大部分は NA および/または DMA にキレートされ、イネのニコチアンミン合成酵素 2 (OsNAS2) 遺伝子の構成的発現 (CE) により白小麦粉中の鉄濃度は最大で 2 倍、亜鉛は最大で 4 倍に増加した。
HMW-TaVIT2 や CE-OsNAS2 のような遺伝子組み換え生物的強化小麦の多地点限定圃場試験(Multi-location confined field trial:CFT)評価は、農作物の性能を維持しながら現実の農業条件下で穀物栄養の増加が維持されるかどうかを判断するために必要である。
私たちは、ガラス温室および圃場条件下で、穀粒の NA、DMA、Fe および Zn をさらに蓄積し、明らかな収量ペナルティを示さない強化CE-OsNAS2 を開発した。ここでは、CE-OsNAS2 小麦についての農学、全粒粉、白粉、白パン栄養について、多地点 CFT で 3 期にわたり包括的に調査した結果について報告する。
(実験方法) 省略
(結果) 省略
(論議)
生物的強化作物開発の重要な要素として、目標とする微量栄養素濃度が、遺伝子型×環境(G×E)相互作用の影響を受けるか、あるいは収量に影響を与えるかを判断するための多地点CFTがある。
以前に、9 つの異なる場所で Zn の生物的強化小麦品種を圃場評価したところ、穀物の Fe および Zn 濃度に有意な G x E 相互作用が見られ、穀物栄養は遺伝子型よりも環境の影響を受けることが確認された。
さらに、OsNAS2 遺伝子とダイズフェリチン(Sfer-H1)遺伝子の胚乳特異的発現を組み合わせた鉄および亜鉛の生物的強化イネ遺伝子導入品種 2 品種の多地点 CFT 評価を行った結果、1 品種(NASFer274)はヌル(null)対照と比較して収量ペナルティがないことが明らかになった。
我々は以前に、CE-1コムギがガラス温室および圃場条件下で栽培された場合、NS植物と明らかな農学的差異を示さないことを報告したが、その研究の圃場要素は、2サイトと1シーズンに限定されていた。
本研究では、3つのCE-1兄弟系統を3つの圃場シーズンにわたって多地点CFTで包括的に評価し、3つのCE-1兄弟系統すべてにおいて、NSと比較して小さいが一貫して有意な草丈の減少を検出した(図1-3)。
S-アデノシルメチオニン(SAM)がエチレンとNA両方の生合成前駆体であり、エチレンが葉の発達、老化、果実の成熟に基本的な役割を果たすことから、NAS遺伝子活性の増加は植物ホルモンのエチレン生合成の変化を介して草丈を減少させると思われる。
エチレンの生合成およびその下流の標的を変更することにより、トウモロコシ (Zea mays, L.) およびイネの作物収量および生物的ストレス耐性が向上することが分かっている。CE-1同胞系統における草丈の減少がエチレン生合成の変化に起因するかどうかはさらなる調査が望まれるところである。しかしながら、3つのCE-1兄弟系統すべてで観察された草丈のわずかな減少は、倒伏の防止に役立つという点で有利である可能性がある。
CE-1 の兄弟系統間の農学的成績、全粒粉栄養および白色小麦粉栄養のばらつき(図 1-3 および補足表 3、5-7)は、体細胞変異によるものと思われる。普及に先立って、CE-1 系統を野生型小麦に戻し交雑してこの変異を取り除く必要があるだろう。
同様の知見はオオムギ (Hordeum vulgare L.) の野外試験評価においても記録されており、そこでは同じ遺伝子導入イベントの兄弟系統間で、第2世代 (T2) および第4世代 (T4) において農学的成績が異なっていた。
これらの結果は、遺伝子組換え植物のCFT評価には、複数世代にわたって行われ、遺伝子組換えイベントごとに複数の兄弟系統を利用すべきであることを示唆している。
重要なことは、本研究では有意な G x E 成分が検出されたが(補足表 2)、圃場間で CE-1 兄弟系統はすべて NS と同等の農業成績と穀物栄養を示したことから、CE-OsNAS2 は圃場条件に関わらず同じ効果を示すことが示唆された。
植物の鉄は主に葉組織にフィトフェリチンとして貯蔵され、登熟期には NA や DMA などの金属キ レーターが葉組織から発達中の穀物への鉄の長距離転流を促進する。登熟期
に CE-1 と NS の葉から穀物に転流する鉄と亜鉛の割合を調べるために、CE-1 と NS の植物体から、ガラス温室(土壌 pH 〜6.0)およびKatanning(土壌 pH 〜4.8)で開化期の止め葉と葉を除去した。止め葉の除去は、CE-1 植物を温室で栽培した場合、NS 植物よりも穀物 Fe および Zn 濃度を低下させたが、Katanning では低下しなかった。このことは、土壌 pH が CE-1 葉から発達中の穀物に微量栄養素の再固定化に影響を与えていることを示している(補 図 5)。
土壌 pH が高いと、土壌中の Fe や Zn の植物への取り込みが制限されるため、土壌 pH の高いガラス温室条件下では、CE-1 株は葉の Fe や Zn をより多く穀物に再固定化できると考えられる。一方、Katanning では土壌 pH が低いため、土壌の Fe および Zn の利用可能性が高くなり(図 1c、補 表 1)、CE-1 の穀粒充填時に地下資源から穀粒への直接再利用がより可能になる可能性がある。
さらに、土壌微量栄養素濃度は、圃場場所間および季節間で異なり(例えば、KatanningとMerredinの土壌Fe濃度は、2016年と2017年に比べて2015年は1.7〜2.4倍高かった)、おそらく最終穀物FeおよびZn濃度に影響を与えるので、多世代にわたるマルチロケーションCFTの実施の重要性がさらに強調されるのである(図1-3)。
CE-1の成熟した止め葉はNSの葉よりも低い(p < 0.05)Feを含んでおり(補足表9)、CE-1植物はNS植物と比較して、登熟時に葉組織から穀物に追加のFeを再固定化することが示唆された。CE-1 の地上部は NS の新芽に比べて NA 濃度が高いことから、この再活性化は CE-1 の葉における NA 濃度の上昇によって媒介されていると思われる30 。
NA は Cu や Mn などの微量栄養素をキレート化することから、CE-1 の葉からの NA を介した再輸送の増加は、CE-1 の全粒粉および白粉中の Cu と Mn 濃度が NS よりも高いことを説明できる(補表 6-8)。
穀粒 Cu 濃度は(白粉 K を除く)他のすべての穀粒栄養素と負の相関を示し、農学的成績形質と正の相関を示したが、NA キレート Cu 濃度を高くする育種が間接的に穀物収量を改善するかどうかはさらなる検討を要する(補足 図 7)。
白粉 K と農学的形質との正の相関は、もともとBobwhite の白粉 K 濃度がチェックに比べて低いことに起因すると思われる(補 表 7)。
NA と DMA は共に S-アデノシルメチオニン(S を含む副基質)から合成される。CE-1 穀物では NA および/または DMA の生合成が促進されることから、すべての CE-1兄弟系統で測定された全粉および白粉の S 濃度が NS と比較して高くなったことが説明できる(補足表6、7)。
本研究では、すべてのCE-1兄弟系統において、全粒粉および白色小麦粉のMg濃度がNSと比較して高いことが確認された。この結果は、OsNAS2 を構成的に過剰発現させたガラス温室栽培小麦でも確認され、NA が Mg2+ イオンをキレートして固定化する能力を有している可能性を示唆するものであった。
今後、CE-1 植物におけるこれらの仮説的な栄養動態を解明し、葉の NA および/または DMA 濃度の増加が小麦の穀物栄養に寄与することを確認するために、放射性同位体およびメタボローム研究が必要である。
Glenthorne、Katanning、Merredin の 3 つの圃場において、我々は CE-1 全粒粉中の Fe、Zn、NA および DMA 濃度を測定し、また CE-1 白粉中の Zn、NA および DMA 濃度を NS と比較したが、これらはすべて有意に高かった(図 1-3)。白粉のFe濃度は、Katanningの3つのフィールドシーズンにおいて、すべてのCE-1兄弟系統で高かった(2017年のCE-1.2を除く)が、GlenthorneとMerredinでは兄弟系統CE-1.1でのみ高かった(p≦0.01)。
FeとZnの両濃度がNSと比較してすべてのCE-1穀粒全体で高いかどうかをさらに明らかにするために、工業規模の製粉(遺伝子型あたり30kg以上)を行い、穀物の最内層と最外層を表す3つのbreak、3つのreduction、pollard、bran(フスマ)の8つの穀物フラクションを得た(表1)。
その結果、CE-1 のbreak、pollard( アリュ−ロン)、およびフスマ画分におい て、NS と比較して有意に高い鉄および亜鉛濃度を検出し、CE-1 粒全体に鉄および亜鉛濃度が高いことを示唆する強い証拠が得られた(表 1、補足表 4)。
また、ガラス温室で栽培した CE-1 および NS 粒子を放射光蛍光 X 線顕微鏡(XFM)分析した結果、CE-1 粒子はアリューロン組織で Fe および Zn が増加し、胚乳組織では Fe シグナルのみが増加した。
これらの結果は、本研究で測定された小麦粉の鉄および亜鉛濃度は、ローラー製粉中に保持される胚乳組織および皺(crease)組織に由来する可能性が高いことを示している。
我々は、すべての CE-1 穀物画分において、NS と比較して有意に高い (p ≤ 0.001) NA および DMA 濃度を検出し、このことはOsNAS2 の構成的発現が小麦粒全体に NA および DMA を増加させることを示唆していた (Table 1)。
破砕(break)および還元(reduction)画分における NA および DMA 濃度の増加は、これらの画分が低濃度のフィチン酸(< 3.5 mg/g)を含み、これらのキレーターが鉄の生物学的利用能を強く向上させることを考えると、おそらく鉄の生物学的利用能を増加させると思われる。
CE-1 外層穀物画分に含まれる高濃度のフィチン酸(> 7.0mg/g)は、in vitro 分析で CE-1 と NS 全粒粉の鉄の生物学的利用能に差がないことが示されたことから、NA および DMA 濃度の増加による鉄の生物学的利用能への影響を相殺すると考えられる。
白色小麦粉の鉄利用率は、本研究を通じてすべてのCE-1同胞系統でNSと比較して高く、白色小麦粉のNAおよびDMA濃度と正の相関があった(図4、補足図6、7)。
我々は以前、鉄濃度に依存しない白粉鉄の生物学的利用能の向上について報告したが、本研究でも同様の結果が兄弟株 CE-1.3 で観察された(図 1g、補足図 6)。これらの結果は、NA および DMA が白小麦粉中の鉄の生物学的利用能を強く向上させ、ヒト細胞への鉄の取り込みを媒介する植物性栄養素であることを示す新たな証拠である。
本研究では、全粒粉および白粉の鉄および亜鉛濃度が穀物収量と負の相関(r ≤ -0.92, p ≤ 0.001)を示し、白粉の NA および DMA 濃度が穀物収量と適度な正の相関(r ; 0.42)を持つことが初めて明らかになった(図 4、補足図 7)。
いくつかの例(例えば、Merredin、2017)では、CE-1兄弟系統は対照に対して穀物NAおよびDMA濃度が高かっただけであり、このことが穀物収量が穀物Fe、Znおよび総タンパク質濃度と負の相関を示し、すべての遺伝子型において穀物NAおよびDMA濃度と相関しなかったことを説明できるかもしれない(図4)。
これらの結果を総合すると、小麦の生物的強化プログラムにおいて、穀物NAおよび/またはDMA濃度を高める育種に重点をシフトすることで、農作物の性能を損なうことなく穀物の両方の鉄の生物的利用性を改善することができることが示唆された。
CE-1 白色パンは、Katanning および Merredin において NS 白色パンと比較して Fe、NA、DMA の濃度が高く、パンのフィチン酸/ Fe 比に差がないことから(補足図 8)、NA および DMA が耐熱性で製パン工程を維持し、 CE-1 白色パンが高い Fe 生物的利用性を示すことが示唆される(図 5)。
全粒粉パンには、フィチン酸、ポリフェノール、食物繊維などの鉄の生物的利用性を阻害する物質が高濃度で含まれていることから、NAとDMA濃度の向上が全粒粉パンの鉄の生物的利用性に及ぼす影響も調べる価値があると思われる。
In vitroスクリーニングツールがないため、CE-1全粒粉および白小麦粉中のZn濃度が一貫して高いことが、Znの生物的利用性を高めることにつながるかどうかを判断することはできない。したがって、CE-1小麦粉とパンの今後の分析には、鉄と亜鉛の生物的利用性を同時に評価する能力を備えた、より包括的なin vivo試験が必要であろう。
白色パンの鉄分濃度は、Glenthorne では NS が CE-1 よりも高く、土壌の性質(特に土壌 pH)が白色パンの鉄分栄養に間接的に影響することが示唆された(図 5a)。Glenthorne の高い土壌 pH は、NS に比べて CE-1 の NA と DMA の生合成を促進し、NA/DMA の Fe に対するモル比を高くしていると思われる。
我々は、Glenthorne の NS 白パンに比べて CE-1 白パンの NA および/または DMA キレート鉄の割合が高いため、crust(パンの皮?)(本研究では採取していない)に比べてcrumb(パンの柔らかい内側)」に局在する鉄の割合が低くなると仮定している。
さらに、CE-1白色小麦粉はNS白色小麦粉に比べてSおよび総タンパク質濃度が高く(補表5および7)、Sおよびタンパク質がグルテン複合体において重要な役割を果たすことから、パン生地のレオロジーに影響を与えると考えられ、CE-1白色パンにおけるFe分布変化にも寄与していると思われる。
今後の実験では、CE-1およびNS白色パンのcrumbクラムとcrustクラストにおけるNAおよび/またはDMAと結合した鉄の局在と空間分布を、放射光ベースの分析によって包括的に明らかにする予定である。
総じて、我々の研究は、後発開発途上国における人間の微量栄養素の摂取量を改善するための、NAバイオ強化した高収量小麦作物(CE-OsNAS2など)の可能性をさらに実証するものである。
以下図の説明。紙面の都合上いくつかの図表は除いている。
Fig.1
イネ由来ニコチアナミン合成酵素2遺伝子を構成的に発現するパンコムギの農学的成績。
(OsNAS2)遺伝子(CE-1.1、CE-1.2、CE-1.3)と、ヌル分離株およびcv. Maceと並べた。
a) T-DNA構築物の模式図。RBとLB:右と左の境界線。
それぞれ;UBI-1:トウモロコシユビキチン1プロモーター;OsNAS2:イネニコチアナミンシンターゼ2遺伝子(LOC_Os03 g19420); NOS: nopaline synthase terminator; 2 x 35S: 35S cauliflower mosaic virus遺伝子のデュアルプロモーター。
hyg: ハイグロマイシンホスホトランスフェラーゼ遺伝子、35S: 35Sカリフラワーモザイクウイルス遺伝子のターミネーター
b) 画像 2017年の圃場シーズンにおけるGlenthorneの反復実験圃場の様子。
c) 土壌pH(緑丸)、および土壌Fe(赤バー)およびZn(濃いグレーのバー)濃度(mg / Kg)は、3つのフィールドシーズンについて記載。
d-j) バーはヌル分離株(ライトグレー)、CE-1.1(ライトブラウン)、CE-1.2(ライトブラウン)の最良線形不偏推定量を示す。
(オレンジ)、CE-1.3 (ダークブラウン)、Mace (黒)のd) 株高 (cm), e) 千粒重 (g), f)
穀物収量(kg ha-1)と,g) Fe (μg / g), h) Zn (μg / g), i) NA (μmol / g), および j) DMA
(μmol / g / g) を全粒粉(左図)と白玉粉(右図)に添加した。各最低3反復
遺伝子型は各フィールドシーズンで含まれ、アスタリスクはNSに対する有意差を表し、p < 0.05 (*), p ; 0.01 (**), p ; 0.001 (***) Wald 検定により決定 n.a = not applicable.
Fig.2
Katanningにおける3つの圃場シーズンでのヌル分離株、CE1.1、CE-1.2、CE-1.3およびMaceの営農成績、全粒および白粉栄養組成
a) 2017年の反復実験圃場シーズンの18m2 のイメージ図。 b) 土壌pH(緑丸)、および土壌Fe(赤棒)、Zn(濃い灰色 )、c-i) 棒グラフは、最良の線形不偏推定量である。
ヌル分離株の推定値(ライトグレー)、CE-1.1(ライトブラウン)、CE-1.2(オレンジ)、CE-1.3(ダークブラウン)、および メイス(黒)については、c) 草丈(cm)、d) 千粒重(g)、e) 穀物収量(kg ha-1)および全粒粉中のf) Fe (μg / g), g) Zn (μg / g), h) NA (μmol / g), i) DMA (μmol / g)の濃度を測定。
(左図)と白玉粉(右図)の2種類がある。各遺伝子型につき最低3つの複製が含まれる。
はNSに対する有意差を表し、p<0.05(*)、p≦0.01(**)、p≦0.001の場合 (***) Wald 検定による。
Fig.5
図5. Glenthorne、Katanning、MerredinにおけるNSおよびCE-1兄弟系統の白色パン栄養組成。
棒グラフは、a) Fe (81;g / g), b) Zn (81;g / g)の濃度に関する最良線形不偏推定量を示している。2016年の各フィールドサイトにおけるc) NA (μmol / g / g), d) DMA (μmol / g/ g)。各遺伝子型につき3連。アスタリスクは p < 0.05 (*), p 0.01 ≤(**) の NS との有意差を表す。Wald 検定により決定した p ≤ 0.001 (***) 。写真は分析前のKatanningから。NSとCE-1ローブの代表的な断面図。 スケールバー=1cm。
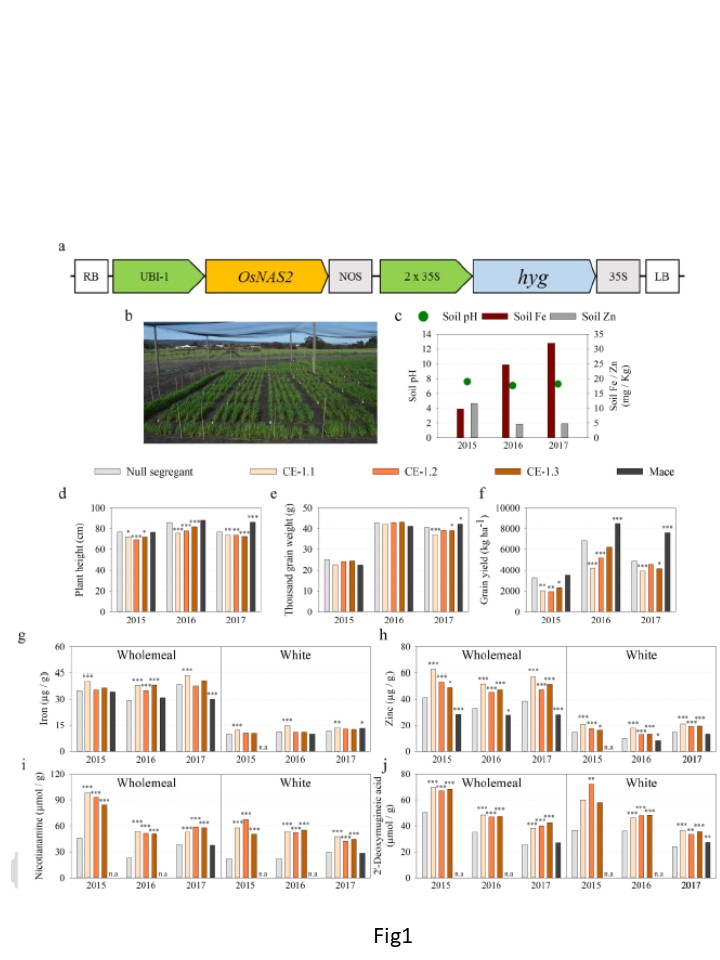
Fig1
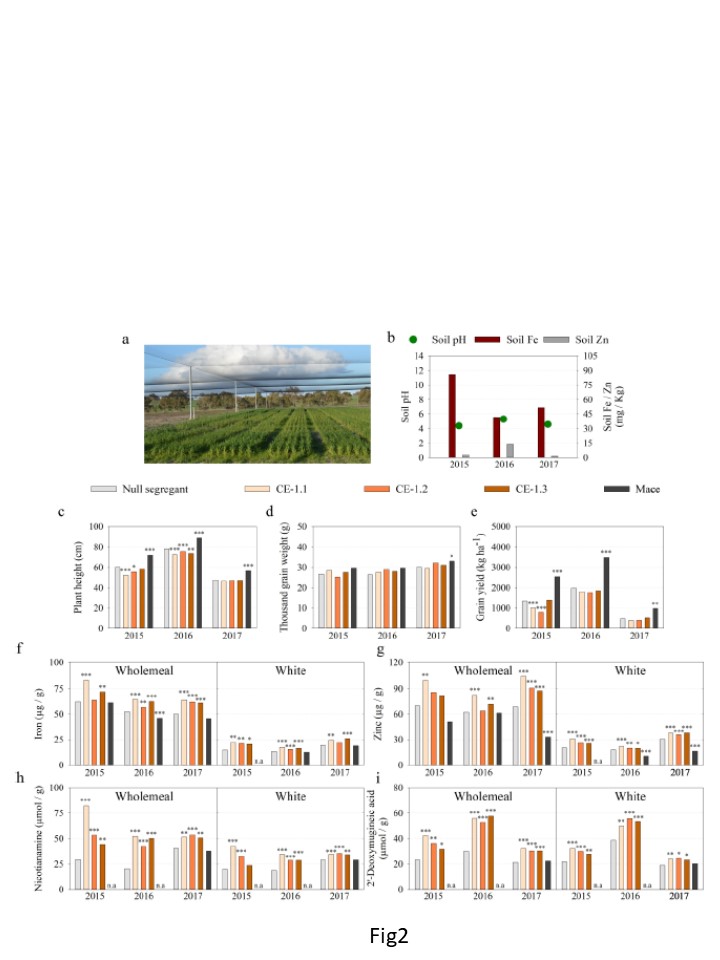
Fig2
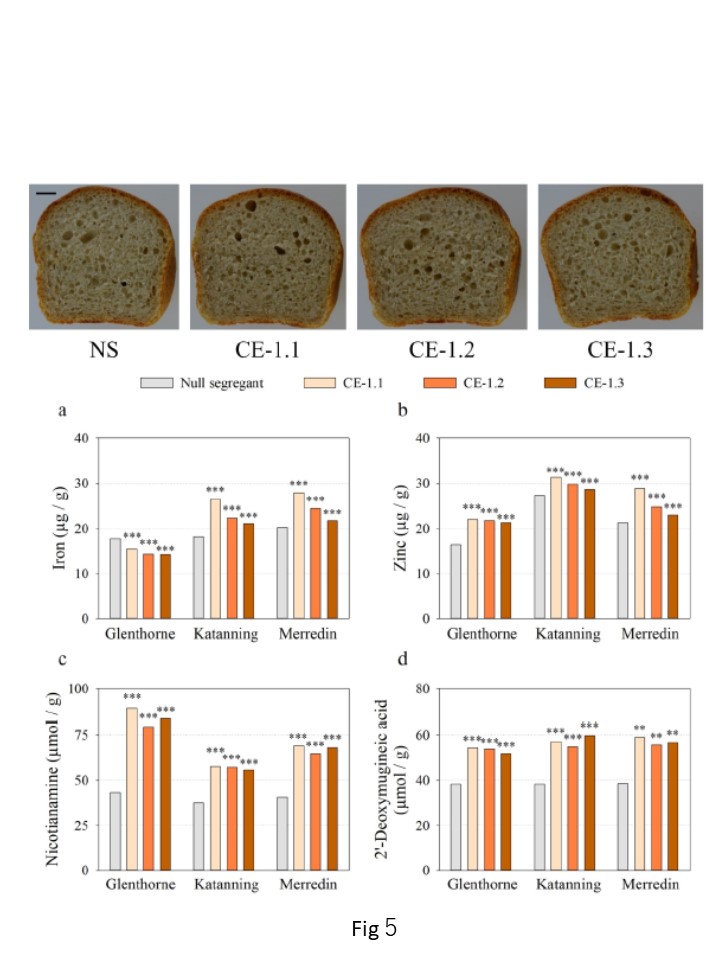
Fig5